
Саркоплазматический ретикулум и гипертрофия: что это и как работает
Саркоплазматический ретикулум — это мембранная система мышечной клетки, которая регулирует концентрацию ионов кальция и запускает механизм сокращения, а гипертрофия представляет собой увеличение объема мышечных волокон за счет роста сократительных белков (миофибриллярный тип) или несократительных структур саркоплазмы (саркоплазматический тип). В покоящейся мышце концентрация свободного кальция составляет менее 10⁻ М, тогда как внутри саркоплазматического ретикулума она достигает 10⁻ М — разница в 10000 раз обеспечивает мгновенный выброс кальция при стимуляции. Понимание механизмов работы ретикулума и типов гипертрофии позволяет целенаправленно управлять тренировочным процессом для развития силы или увеличения мышечной массы.
Что такое саркоплазматический ретикулум и зачем он мышечной клетке
Саркоплазматический ретикулум (СР) — это специализированная мембранная органелла мышечных клеток, главная функция которой заключается в хранении и контролируемом высвобождении ионов кальция для запуска мышечного сокращения. Это своего рода внутриклеточный резервуар, который окружает каждую сократительную единицу мышцы подобно кружевной манжете.
Открытие этой структуры произошло в 1901 году, когда итальянский гистолог Эмилио Вератти впервые обнаружил тончайшую сеть в мышечном волокне с помощью специальных методов окрашивания. Однако полное понимание строения и функций саркоплазматического ретикулума стало возможным только в середине XX века с изобретением электронного микроскопа, который позволил увидеть детали структуры размером до нескольких нанометров.
Концентрация ионов кальция внутри мышечной клетки в состоянии покоя поддерживается в 100 000 раз ниже, чем во внеклеточной жидкости. Этот огромный градиент концентрации создает саркоплазматический ретикулум, что позволяет мгновенно увеличивать уровень кальция в цитоплазме при необходимости сокращения.
Основная биологическая роль саркоплазматического ретикулума состоит в решении фундаментальной задачи: обеспечении быстрого и контролируемого перехода мышцы из состояния покоя в состояние сокращения и обратно. Чтобы понять важность этой системы, можно провести аналогию с гидравлическим прессом. Представьте, что кальций — это рабочая жидкость, которая приводит в действие механизм сокращения. СР работает как резервуар высокого давления, который может мгновенно выпустить эту жидкость в нужный момент, а затем быстро откачать ее обратно, возвращая систему в исходное состояние.
В покоящейся мышечной клетке концентрация свободных ионов кальция в саркоплазме (цитоплазме мышечной клетки) составляет менее 10-7 моль на литр (М). В то же время внутри саркоплазматического ретикулума концентрация кальция достигает 10-3 М — разница составляет 10 000 раз. Эта разница не случайна: слишком высокий уровень кальция в цитоплазме токсичен для клетки, поскольку приводит к кальцификации митохондрий и других внутриклеточных структур, что в конечном итоге вызывает гибель клетки.
Почему именно кальций используется для запуска сокращения
Ионы кальция обладают уникальными химическими свойствами, которые делают их идеальными сигнальными молекулами в клетке. Во-первых, кальций имеет два положительных заряда, что позволяет ему образовывать прочные связи с белками. Во-вторых, ионный радиус кальция обеспечивает высокую избирательность связывания с определенными белками. В-третьих, клетка может легко создать значительный градиент концентрации кальция между внутренним и внешним пространством, что критично для быстрой сигнализации. Альтернативные ионы, такие как магний, не обладают таким сочетанием свойств для выполнения роли вторичного посредника в мышечном сокращении.
Саркоплазматический ретикулум выполняет три критические функции в мышечной клетке. Первая — активное накопление и концентрирование ионов кальция из цитоплазмы с помощью специальных кальциевых насосов (Ca2+-АТФаз), которые используют энергию АТФ. Вторая — хранение накопленного кальция в связанном состоянии с помощью специального белка кальсеквестрина, который может связывать до 50 ионов кальция на одну молекулу благодаря высокому содержанию отрицательно заряженных аминокислот. Третья — контролируемое высвобождение кальция в ответ на электрический сигнал, поступающий от нервной системы.
Цена отсутствия или повреждения саркоплазматического ретикулума крайне высока. При повреждении мембран СР происходит неконтролируемый выброс кальция в саркоплазму, что активирует ферменты-протеазы, разрушающие мышечные белки. Это один из механизмов повреждения мышечных волокон при интенсивных эксцентрических нагрузках или при некоторых мышечных дистрофиях. Восстановление поврежденного СР требует синтеза новых мембранных компонентов и белков, что занимает от 48 до 72 часов — именно поэтому отсроченная мышечная боль после тренировки может длиться несколько дней.
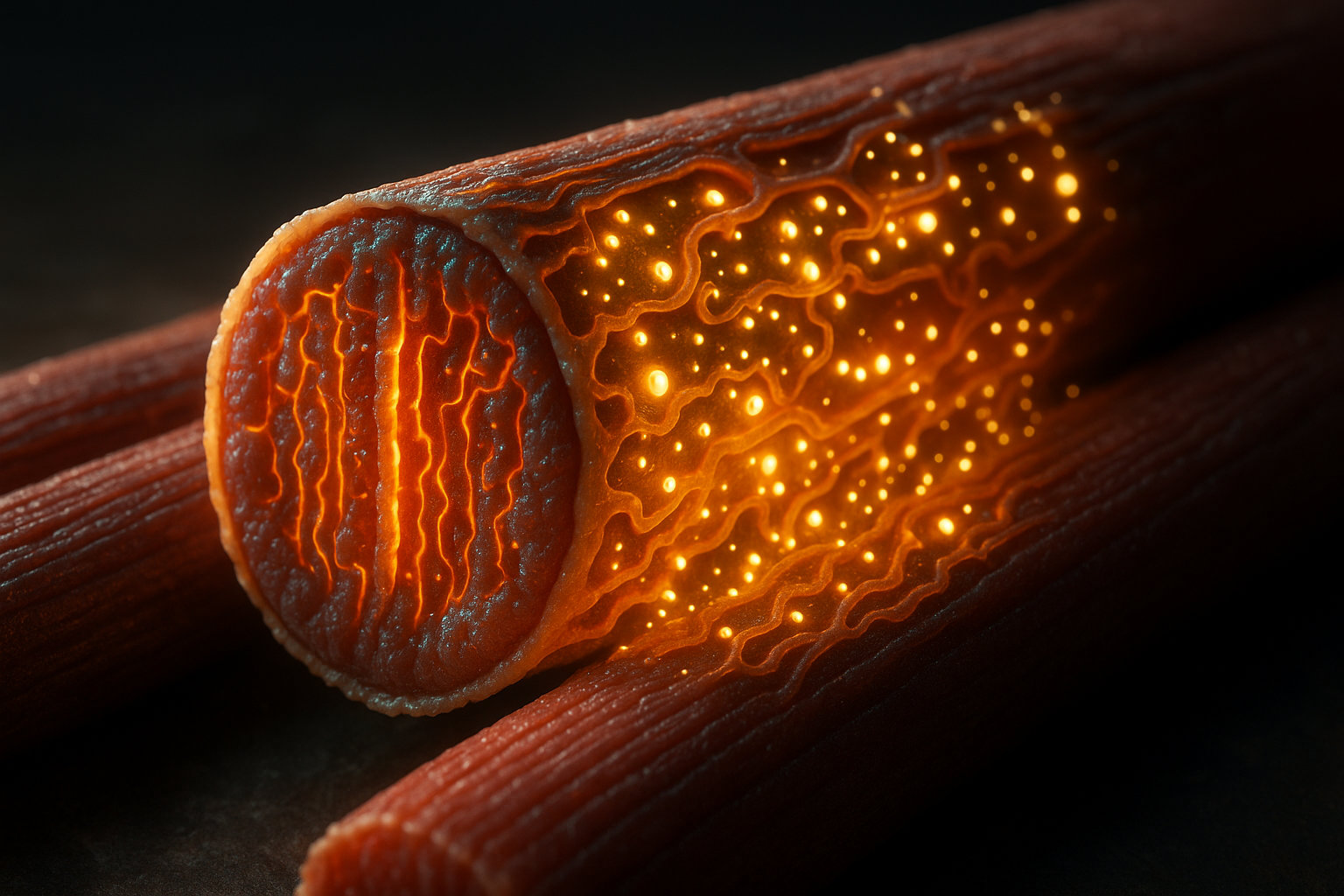
Структура саркоплазматического ретикулума: от терминальных цистерн до T-трубочек
Саркоплазматический ретикулум представляет собой трехмерную сеть взаимосвязанных мембранных трубочек и мешочков, которые опутывают каждую миофибриллу мышечного волокна. Архитектура этой системы напоминает кружевной чулок, натянутый на сократительный аппарат мышцы, где каждая миофибрилла имеет свою собственную оболочку из СР.
Структурно саркоплазматический ретикулум делится на два функционально различных компонента. Продольные элементы (L-канальцы) — это тонкие трубочки диаметром 30-60 нанометров, которые идут параллельно миофибриллам и соединяются между собой, образуя непрерывную сеть. Эти элементы занимают основной объем СР и содержат высокую концентрацию кальциевых насосов (Ca2+-АТФаз), которые активно закачивают кальций обратно в ретикулум после сокращения. Второй компонент — терминальные цистерны (конечные цистерны) — это расширенные участки ретикулума диаметром до 100-200 нанометров, которые располагаются в строго определенных местах вдоль миофибриллы.
| Компонент структуры | Размер | Локализация | Основная функция |
|---|---|---|---|
| Продольные элементы (L-канальцы) | 30-60 нм | Вдоль миофибрилл | Обратный захват Ca2+ из цитоплазмы |
| Терминальные цистерны | 100-200 нм | В области Z-дисков (сердечная мышца) или на границе A-I полос (скелетная мышца) | Хранение и высвобождение Ca2+ |
| T-трубочки | 20-450 нм | Перпендикулярно миофибриллам | Проведение электрического сигнала вглубь волокна |
Терминальные цистерны содержат внутри себя белок кальсеквестрин в высокой концентрации (до 40 миллимоль на литр внутреннего объема), что позволяет хранить огромное количество ионов кальция в относительно небольшом пространстве. На мембране цистерн располагаются рианодиновые рецепторы — крупные белковые комплексы, которые формируют кальциевые каналы и служат точками выхода кальция в саркоплазму при активации.
Критически важным элементом архитектуры является система T-трубочек (поперечных трубочек) — это не часть саркоплазматического ретикулума, а впячивания наружной клеточной мембраны (сарколеммы), которые проникают вглубь мышечного волокна. T-трубочки были впервые описаны в 1897 году, когда исследователи ввели чернила в сердечную мышцу и обнаружили, что красящее вещество проникает внутрь клетки по определенным каналам. Эти структуры располагаются перпендикулярно или под углом к миофибриллам и доходят до самого центра мышечного волокна.
Скорость проведения потенциала действия вдоль T-трубочек составляет около 2 метров в секунду, что позволяет электрическому сигналу достичь центральных миофибрилл мышечного волокна диаметром 100 микрометров менее чем за 50 микросекунд.
Терминальные цистерны саркоплазматического ретикулума и T-трубочки образуют функциональные комплексы, которые различаются в зависимости от типа мышцы. В скелетных мышцах формируются триады — структуры, где одна T-трубочка расположена между двумя терминальными цистернами СР. Расстояние между мембраной T-трубочки и мембраной цистерны составляет всего 12-15 нанометров — это критично для быстрого взаимодействия белков. В сердечной мышце обычно формируются диады — комплексы из одной T-трубочки и одной терминальной цистерны.
Мембрана T-трубочек насыщена специализированными белками: кальциевыми каналами L-типа, натрий-кальциевыми обменниками, кальциевыми АТФазами и β-адренорецепторами. Концентрация кальциевых каналов L-типа в T-трубочках в 3-5 раз выше, чем в остальной части клеточной мембраны. Эта высокая плотность каналов обеспечивает эффективное электромеханическое сопряжение — процесс преобразования электрического сигнала в механическое сокращение.
Различия в организации триад и диад: компромисс между скоростью и точностью
Скелетные мышцы используют триадную организацию, где кальциевые каналы L-типа на T-трубочке физически связаны с рианодиновыми рецепторами на СР через белковые мостики. Это обеспечивает сверхбыструю активацию без необходимости входа кальция извне — механическое взаимодействие белков запускает открытие рианодиновых рецепторов напрямую. Цена этого решения — меньшая гибкость регуляции. Сердечная мышца использует диады с механизмом кальций-индуцированного выхода кальция: небольшое количество кальция входит через L-каналы и запускает массивный выброс из СР. Это медленнее на 5-10 миллисекунд, но позволяет тонко регулировать силу сокращения в зависимости от потребностей организма.
Структурная организация СР не статична — она адаптируется к функциональным требованиям. При регулярных тренировках на выносливость объем саркоплазматического ретикулума может увеличиваться на 15-25% за счет синтеза дополнительных мембран и белков. Это увеличение коррелирует с повышением способности мышцы к повторным сокращениям, поскольку больший объем СР означает более эффективное удаление кальция из саркоплазмы и более быстрое расслабление мышцы между сокращениями.

Как саркоплазматический ретикулум управляет концентрацией кальция
Управление концентрацией кальция саркоплазматическим ретикулумом представляет собой двухфазный процесс: быструю фазу высвобождения и медленную фазу обратного захвата. Система работает как прецизионный дозатор, способный за миллисекунды изменить концентрацию кальция в саркоплазме в 100-1000 раз и затем вернуть ее к исходному уровню.
Процесс высвобождения кальция начинается с прихода потенциала действия по T-трубочке. Деполяризация мембраны T-трубочки активирует кальциевые каналы L-типа (дигидропиридиновые рецепторы), которые изменяют свою конформацию в течение 1-2 миллисекунд. В скелетных мышцах эти каналы механически связаны с рианодиновыми рецепторами на мембране терминальных цистерн через специальные белковые мостики. Конформационное изменение L-канала передается непосредственно на рианодиновый рецептор, открывая его без необходимости входа кальция извне.
Открытие рианодиновых рецепторов запускает массивный выброс ионов кальция из терминальных цистерн. Скорость выхода кальция через один открытый канал составляет около 10 миллионов ионов в секунду. Учитывая, что в одной терминальной цистерне может находиться несколько сотен рианодиновых рецепторов, суммарный поток кальция достигает колоссальных значений. За 5-10 миллисекунд концентрация свободного кальция в пространстве между тонкими и толстыми филаментами миофибриллы возрастает с базовых 10-7 М до пиковых 10-5 - 10-4 М — увеличение в 100-1000 раз.
Саркоплазматический ретикулум скелетной мышцы содержит около 90% всех мембранных Ca2+-АТФаз в организме. Эти молекулярные насосы способны переносить до 70% всей клеточной энергии АТФ в высокоактивных мышечных волокнах во время интенсивной работы.
Обратный захват кальция из саркоплазмы обратно в саркоплазматический ретикулум осуществляется Ca2+-АТФазами саркоплазматического ретикулума (SERCA — Sarco/Endoplasmic Reticulum Ca2+-ATPase). Эти молекулярные насосы составляют до 80% всех мембранных белков продольных элементов СР — самая высокая концентрация одного типа белка среди всех известных биологических мембран. Каждый насос способен транспортировать 2 иона кальция за один цикл работы, расходуя при этом одну молекулу АТФ.
Механизм работы SERCA включает несколько конформационных состояний. На стороне, обращенной к саркоплазме, насос связывает два иона кальция с высоким сродством. Гидролиз АТФ приводит к фосфорилированию белка и изменению его конформации, при котором кальций-связывающие сайты переориентируются в просвет СР, а сродство к кальцию снижается. Ионы кальция высвобождаются внутрь ретикулума, где немедленно связываются с кальсеквестрином. Дефосфорилирование белка возвращает его в исходную конформацию, готовую к новому циклу. Полный цикл занимает около 10-20 миллисекунд при комнатной температуре и 5-10 миллисекунд при температуре тела человека.
| Этап процесса | Ключевые белки | Временная шкала | Энергетическая цена |
|---|---|---|---|
| Высвобождение Ca2+ | Рианодиновые рецепторы (RyR) | 5-10 миллисекунд | Не требует АТФ (пассивный процесс) |
| Обратный захват Ca2+ | SERCA-насосы | 50-200 миллисекунд | 1 АТФ на 2 иона Ca2+ |
| Хранение Ca2+ в СР | Кальсеквестрин | Постоянно | Не требует энергии |
Регуляция активности SERCA осуществляется небольшим белком фосфоламбаном, который в дефосфорилированном состоянии ингибирует насос, снижая его сродство к кальцию. При стимуляции β-адренорецепторов (например, адреналином во время стресса) фосфоламбан фосфорилируется протеинкиназой А, что снимает ингибирование и увеличивает скорость обратного захвата кальция в 2-3 раза. Этот механизм объясняет, почему сердце бьется быстрее и сильнее при выбросе адреналина — ускорение удаления кальция позволяет мышце быстрее расслабляться между сокращениями.
Эффективность системы контроля кальция определяет физиологические возможности мышцы. В быстрых гликолитических волокнах (тип IIx) плотность SERCA-насосов на 40-60% выше, чем в медленных окислительных волокнах (тип I), что обеспечивает быстрое расслабление и возможность работать с высокой частотой сокращений. Однако эта адаптация имеет цену: высокая активность SERCA потребляет огромное количество АТФ. При интенсивной работе быстрых волокон до 30-40% всей произведенной АТФ расходуется исключительно на работу кальциевых насосов, а не на само сокращение.
Почему мышца не может сокращаться бесконечно быстро
Теоретический предел частоты сокращений определяется скоростью работы SERCA-насосов. Даже если нервная система посылает импульсы с очень высокой частотой, мышца не сможет полностью расслабиться между сокращениями, если кальций не будет удален из саркоплазмы. У человека максимальная частота полностью разделенных сокращений составляет около 10-15 сокращений в секунду для самых быстрых мышц. При более высоких частотах стимуляции происходит суммация сокращений и развивается тетанус — состояние непрерывного сокращения. Интересно, что у некоторых специализированных мышц, например, у певчих птиц (syrinx) или у гремучих змей (мышцы погремушки), эволюция создала экстремально высокую плотность SERCA и модифицированные изоформы этого фермента, позволяющие достигать частоты до 100-200 сокращений в секунду.
При физической тренировке адаптация системы контроля кальция происходит по-разному в зависимости от типа нагрузки. Тренировки на выносливость увеличивают общий объем саркоплазматического ретикулума на 15-25% и повышают экспрессию SERCA, что улучшает способность мышцы к длительной работе. Силовые тренировки с максимальными весами меньше влияют на плотность SERCA, но могут изменять соотношение изоформ этого фермента в сторону более быстрых вариантов, что ускоряет релаксацию между повторениями с тяжелыми весами.
Роль ионов кальция в механизме мышечного сокращения
Ионы кальция выполняют функцию молекулярного переключателя, который запускает процесс сокращения мышцы: при повышении концентрации Ca2+ с 10-7 М до 10-5 М кальций связывается с белком тропонином С, что снимает блокировку с активных центров актина и позволяет головкам миозина прикрепиться к нему. Без кальция мышечное сокращение физически невозможно, даже если в клетке есть достаточно энергии в виде АТФ.
В состоянии покоя тонкие актиновые филаменты мышечного волокна блокированы для взаимодействия с миозином специальным белковым комплексом. Тропомиозин — длинная белковая нить, которая располагается в желобке между двумя спиральными цепями актина и физически закрывает участки связывания миозина. Этот механизм можно сравнить с защитной крышкой на розетке — пока крышка закрыта, контакт невозможен, даже если вилка находится рядом.
На тропомиозине через каждые 38,5 нанометров располагается еще один белок — тропонин, который состоит из трех субъединиц с различными функциями. Тропонин Т (TnT) прикрепляет весь комплекс к тропомиозину, тропонин I (TnI) в отсутствие кальция усиливает связывание тропомиозина с актином и блокирует миозин-связывающие сайты, а тропонин С (TnC) содержит четыре участка связывания ионов кальция и служит кальциевым сенсором системы.
При концентрации свободного кальция 10-7 М мышечное волокно генерирует нулевую силу. При повышении до 10-6 М сила сокращения составляет около 50% от максимальной, а при 10-5 М достигается 100% силы. Такая чувствительность позволяет тонко регулировать силу сокращения изменением концентрации кальция всего в 10-100 раз.
Когда ионы кальция выходят из саркоплазматического ретикулума, они связываются с тропонином С в течение 1-2 миллисекунд. Присоединение двух ионов кальция к TnC вызывает конформационные изменения во всем тропонин-тропомиозиновом комплексе. Тропонин I ослабляет свою связь с актином, а тропомиозин смещается глубже в желобок между актиновыми цепями — примерно на 1-1,5 нанометра. Этого небольшого смещения достаточно, чтобы открыть участки связывания для головок миозина.
После открытия активных центров начинается циклическое взаимодействие миозина с актином, известное как перекрестный мостиковый цикл. Головка миозина, на которой уже находится АТФ в гидролизованной форме (АДФ и неорганический фосфат), прикрепляется к открытому участку актина. Это вызывает высвобождение фосфата и конформационное изменение головки миозина — рабочий ход, при котором актиновый филамент смещается относительно миозинового на 10-12 нанометров. После этого отсоединяется АДФ, и на его место связывается новая молекула АТФ, что заставляет головку миозина отсоединиться от актина.
| Этап цикла | Конформация миозина | Связанные нуклеотиды | Длительность (мс) |
|---|---|---|---|
| Присоединение к актину | Слабосвязанное состояние | АДФ + Pi (фосфат) | 1-2 |
| Рабочий ход (смещение актина) | Сильносвязанное состояние | АДФ (после высвобождения Pi) | 5-10 |
| Отсоединение от актина | Несвязанное состояние | АТФ | 0,1-0,5 |
| Гидролиз АТФ и возврат | Готовность к новому циклу | АДФ + Pi | 2-5 |
Один полный цикл взаимодействия миозина с актином при температуре тела человека занимает около 10-20 миллисекунд в быстрых мышечных волокнах и 50-100 миллисекунд в медленных волокнах. За это время генерируется сила примерно 3-5 пиконьютонов на одну миозиновую головку. Учитывая, что в одном толстом филаменте содержится около 300 молекул миозина (600 головок), а в одной миофибрилле — тысячи толстых филаментов, суммарная сила становится достаточной для движения конечностей и перемещения тела.
Почему для расслабления мышцы нужна энергия АТФ
Распространенное заблуждение состоит в том, что энергия нужна только для сокращения мышцы. В действительности для отсоединения миозина от актина также требуется АТФ. В отсутствие АТФ головки миозина остаются прикрепленными к актину в так называемом состоянии rigor (от лат. "жесткость"). Именно это состояние наблюдается при трупном окоченении (rigor mortis): после смерти прекращается синтез АТФ, существующие запасы быстро истощаются, и мышцы застывают в жестком состоянии. Через 48-72 часа начинается распад белков, и окоченение проходит. Этот факт подчеркивает, что АТФ нужна не только для совершения работы (рабочего хода миозина), но и для разъединения актин-миозинового комплекса, что позволяет мышце расслабиться.
Регуляция чувствительности мышечных волокон к кальцию может изменяться в зависимости от физиологических условий. При снижении pH (накоплении молочной кислоты во время интенсивной работы) сродство тропонина С к кальцию уменьшается на 20-30%, что означает необходимость более высокой концентрации кальция для достижения той же силы сокращения. Это один из механизмов мышечного утомления при высокоинтенсивных нагрузках. Температура также влияет на систему: при охлаждении мышцы на 10°C скорость присоединения и отсоединения кальция от тропонина замедляется примерно в 2-3 раза, что объясняет снижение скорости сокращения холодных мышц.
Интересно, что эволюция создала два разных механизма кальциевой регуляции мышечного сокращения. У позвоночных животных, как описано выше, кальций регулирует сокращение через тонкие актиновые филаменты и тропонин. У большинства беспозвоночных (моллюсков, членистоногих) кальций действует непосредственно на толстые миозиновые филаменты, связываясь с легкими цепями миозина и активируя их. Оба механизма эффективны, но позвоночный вариант обеспечивает более быструю и точную регуляцию благодаря кооперативному эффекту: один тропонин-тропомиозиновый комплекс контролирует сразу семь миозин-связывающих участков на актине.

Почему концентрация кальция в клетке поддерживается в 100000 раз ниже внеклеточной
Концентрация свободного кальция в цитоплазме покоящейся мышечной клетки составляет 10-7 - 10-8 М, тогда как во внеклеточной жидкости она достигает 10-3 М — разница в 10000-100000 раз создает мощный электрохимический градиент, который используется клеткой для быстрой сигнализации. Поддержание низкой концентрации кальция внутри клетки критично для выживания, поскольку избыток кальция запускает каскад токсических реакций, ведущих к гибели клетки.
Основная причина токсичности высоких концентраций кальция связана с тем, что этот ион активирует множество ферментов, которые в нормальных условиях должны находиться в неактивном состоянии. При концентрации выше 10-6 М кальций активирует кальпаины — протеазы, разрушающие цитоскелетные и сократительные белки клетки. Эти ферменты начинают расщеплять белки, необходимые для структурной целостности клетки, что приводит к ее разрушению.
Кроме того, избыточный кальций накапливается в митохондриях — энергетических станциях клетки. Митохондрии обладают способностью поглощать кальций из цитоплазмы, что в норме служит буферной системой. Однако когда концентрация кальция в митохондриях превышает пороговое значение (около 1-2 миллимоль на литр митохондриального матрикса), происходит открытие митохондриальной поры переходной проницаемости. Через эту пору из митохондрий выходят белки, запускающие программу клеточной смерти (апоптоз), и прекращается синтез АТФ.
Экспериментальные данные показывают, что если в мышечной клетке концентрация свободного кальция искусственно поддерживается на уровне 10-5 М (что соответствует пиковому значению при сокращении) в течение более 30 минут, начинается необратимое повреждение клетки. При 10-4 М клетка погибает в течение 5-10 минут.
Огромный градиент концентрации кальция между внешней средой и цитоплазмой создает естественную систему усиления сигнала. Представьте это как плотину, за которой накопилась вода под высоким давлением. Достаточно открыть небольшую заслонку, и мощный поток воды устремится вниз. Аналогично, открытие даже небольшого количества кальциевых каналов в мембране саркоплазматического ретикулума приводит к массивному входу кальция в цитоплазму благодаря огромной разнице концентраций.
Для поддержания низкой концентрации кальция в цитоплазме клетка использует несколько энергозависимых систем. Основную работу выполняют SERCA-насосы саркоплазматического ретикулума, о которых говорилось ранее. Дополнительно работает плазмалеммальная Ca2+-АТФаза (PMCA), которая выкачивает кальций из клетки наружу через поверхностную мембрану. Третья система — натрий-кальциевый обменник (NCX), который использует энергию натриевого градиента для выведения одного иона кальция в обмен на вход трех ионов натрия.
| Система транспорта Ca2+ | Локализация | Скорость работы | Вклад в удаление Ca2+ | Энергетическая цена |
|---|---|---|---|---|
| SERCA-насосы | Мембрана СР | Высокая (100-200 ионов/с на насос) | 70-90% в скелетных мышцах | 1 АТФ на 2 Ca2+ |
| PMCA-насосы | Плазматическая мембрана | Низкая (30-40 ионов/с на насос) | 5-10% | 1 АТФ на 1 Ca2+ |
| NCX-обменник | Плазматическая мембрана | Очень высокая (>1000 ионов/с) | 10-20% (больше в сердце) | Косвенная (через Na+-K+-АТФазу) |
| Митохондрии | Внутри клетки | Переменная | 5-10% (буферная функция) | Не требует АТФ напрямую |
Энергетическая цена поддержания кальциевого градиента огромна. В покоящейся мышце на работу кальциевых насосов тратится около 10-15% всей производимой АТФ, несмотря на то что мышца не сокращается. Во время активной работы эта доля возрастает до 30-40%. Это означает, что примерно треть всей энергии, которую мышца тратит при интенсивном сокращении, идет не на выполнение механической работы, а на поддержание кальциевого гомеостаза.
Компромисс данной системы заключается в следующем: с одной стороны, огромный градиент обеспечивает быстроту и мощность сигнала (мышца может сократиться за миллисекунды), с другой стороны, поддержание этого градиента требует постоянных энергетических затрат даже в покое. Эволюция выбрала этот путь, поскольку способность быстро реагировать на опасность критична для выживания — цена медленной реакции (быть съеденным хищником) намного выше, чем цена постоянных энергетических расходов на кальциевые насосы.
Альтернативные стратегии сигнализации у других организмов
Интересно, что не все организмы используют столь энергозатратную систему кальциевой сигнализации. Некоторые примитивные многоклеточные организмы используют другие ионы для запуска быстрых ответов. Например, у некоторых растений аналогичную роль играют ионы хлора или калия. Однако в эволюции животных кальциевая сигнализация стала доминирующей, вероятно, из-за уникальной способности кальция образовывать высокоспецифичные связи с белками и создавать устойчивые градиенты благодаря низкой растворимости солей кальция с фосфатами и карбонатами, которые могут служить внутриклеточными буферами.
При старении и некоторых патологических состояниях способность клетки поддерживать низкую концентрацию кальция снижается. Экспрессия SERCA-насосов может уменьшаться на 20-40%, что приводит к более медленному расслаблению мышц и снижению их работоспособности. Это один из механизмов возрастного снижения мышечной функции. Регулярные физические тренировки умеренной интенсивности помогают поддерживать экспрессию кальциевых насосов на высоком уровне, что частично компенсирует возрастные изменения.
Что происходит при повреждении саркоплазматического ретикулума
Повреждение мембран саркоплазматического ретикулума приводит к неконтролируемой утечке ионов кальция в цитоплазму и запускает каскад разрушительных процессов в мышечной клетке: концентрация Ca2+ может оставаться повышенной в течение часов вместо миллисекунд, что активирует протеолитические ферменты, разрушающие сократительные белки и клеточные структуры. Это основной механизм повреждения мышц при интенсивных эксцентрических нагрузках, приводящий к отсроченной мышечной боли и временному снижению силовых показателей.
Наиболее уязвимым компонентом саркоплазматического ретикулума являются терминальные цистерны — расширенные участки, где хранится основной запас кальция. При механическом растяжении мышцы во время эксцентрического сокращения (когда мышца удлиняется под нагрузкой, например, при опускании тяжелого веса) мембраны цистерн испытывают повышенное напряжение. Если напряжение превышает прочность мембраны, образуются микроразрывы размером от нескольких до десятков нанометров.
Через эти разрывы кальций начинает поступать в саркоплазму со скоростью, которая превышает возможности SERCA-насосов по его удалению. В результате концентрация свободного кальция в поврежденной области может подняться до 10-5 - 10-4 М и оставаться на этом уровне в течение 30-60 минут вместо нормальных 50-200 миллисекунд. Это кальциевая перегрузка запускает несколько параллельных деструктивных процессов.
Эксперименты с изолированными мышечными волокнами показывают, что повреждение саркоплазматического ретикулума приводит к снижению максимальной силы сокращения на 30-50% в течение первых 24-48 часов. Полное восстановление функции может занимать от 5 до 10 дней в зависимости от степени повреждения.
Первая линия разрушения — активация кальпаинов, кальций-зависимых протеаз, которые разрушают цитоскелетные белки. Кальпаины разрезают белки, соединяющие миофибриллы с сарколеммой (титин, десмин, дистрофин), что нарушает передачу силы от сократительного аппарата к сухожилиям. Особенно страдают Z-диски — структуры, к которым прикрепляются тонкие актиновые филаменты. Электронная микроскопия поврежденных мышц показывает характерную картину размытых, расплывчатых Z-дисков, получившую название Z-линейный стриминг.
Вторая линия повреждения связана с митохондриями. Избыточный кальций поглощается митохондриями, которые пытаются выполнять буферную функцию. Однако их емкость ограничена, и при перегрузке происходит открытие митохондриальной поры переходной проницаемости. Это приводит к набуханию митохондрий, разрушению их мембран и высвобождению проапоптотических факторов. Клетка теряет способность производить достаточное количество АТФ, что еще больше нарушает работу кальциевых насосов и создает порочный круг.
| Стадия повреждения | Временные рамки | Основные процессы | Клинические проявления |
|---|---|---|---|
| Острая фаза | 0-6 часов после нагрузки | Разрыв мембран СР, утечка Ca2+, активация кальпаинов | Минимальная боль, небольшое снижение силы (10-20%) |
| Воспалительная фаза | 6-48 часов | Инфильтрация нейтрофилов, выработка свободных радикалов, отек | Отсроченная мышечная боль (DOMS), снижение силы на 30-50% |
| Регенеративная фаза | 2-5 дней | Активация сателлитных клеток, синтез новых белков СР | Постепенное снижение боли, восстановление силы |
| Ремоделирование | 5-14 дней | Укрепление структур, адаптация к нагрузке | Полное восстановление функции, эффект повторной тренировки (repeated bout effect) |
Третий механизм повреждения — воспалительная реакция. Поврежденные мышечные волокна выделяют сигнальные молекулы (цитокины, хемокины), которые привлекают клетки иммунной системы — нейтрофилы и макрофаги. Эти клетки проникают в мышечную ткань в течение 6-24 часов после повреждения и начинают вырабатывать активные формы кислорода и протеолитические ферменты для очистки поврежденных структур. Именно эта воспалительная фаза является основной причиной отсроченной мышечной боли (DOMS — Delayed Onset Muscle Soreness), которая достигает пика через 24-48 часов после тренировки.
Интересный феномен, связанный с повреждением саркоплазматического ретикулума, — эффект повторной тренировки (repeated bout effect). После первого эпизода повреждающей нагрузки мышца становится более устойчивой к последующим аналогичным нагрузкам в течение 6-8 недель. Механизмы этой адаптации включают увеличение прочности мембран СР за счет изменения липидного состава, повышение экспрессии белков теплового шока (HSP), которые защищают клеточные структуры от повреждений, и улучшение системы репарации мембран.
Практические рекомендации для минимизации повреждений СР при тренировках
Чтобы снизить риск серьезного повреждения саркоплазматического ретикулума при тренировках, особенно при освоении новых упражнений или возвращении после перерыва, рекомендуется использовать принцип постепенной прогрессии. В первой тренировке с новым движением или после длительного перерыва используйте нагрузку не более 50-60% от максимальной и ограничьте объем эксцентрических повторений. Через 7-10 дней, когда пройдет адаптация, можно безопасно увеличивать нагрузку. Адекватное потребление белка (1,6-2,2 г на кг массы тела) и достаточный сон (7-9 часов) критичны для восстановления поврежденных структур. Прием антиоксидантов в высоких дозах непосредственно после тренировки может парадоксально замедлить адаптацию, поскольку умеренный окислительный стресс служит сигналом для запуска восстановительных процессов.
В некоторых патологических состояниях повреждение саркоплазматического ретикулума становится хроническим. При мышечных дистрофиях (например, дистрофии Дюшенна) генетические дефекты белков, связывающих сарколемму с цитоскелетом, приводят к повторяющимся разрывам мембран при каждом сокращении. Это вызывает постоянную кальциевую перегрузку и прогрессирующую дегенерацию мышечных волокон. При злокачественной гипертермии — редком наследственном заболевании — мутация рианодиновых рецепторов приводит к их спонтанному открытию при воздействии некоторых анестетиков, что вызывает массивный и неконтролируемый выброс кальция из СР, тяжелое повреждение мышц и может привести к летальному исходу без экстренного лечения.
Восстановление поврежденного саркоплазматического ретикулума требует синтеза новых мембранных липидов и белков, что является энергозатратным процессом. Клетка активирует экспрессию генов, кодирующих SERCA, рианодиновые рецепторы, кальсеквестрин и другие компоненты СР. Скорость синтеза составляет примерно 2-5% от общего пула белков СР в сутки, что объясняет, почему полное функциональное восстановление после серьезного повреждения занимает от 5 до 10 дней.
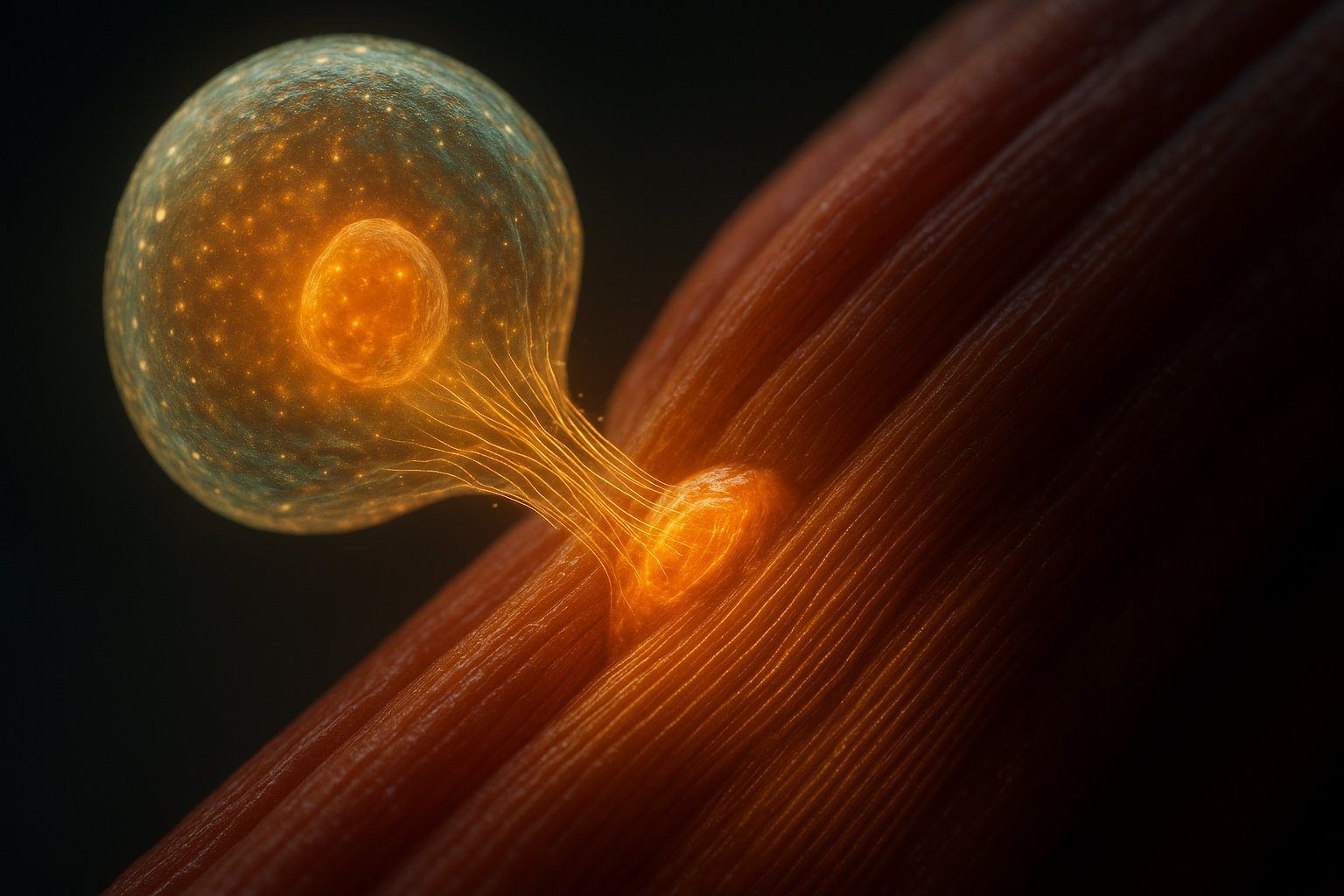
Гипертрофия скелетных мышц: определение и базовые механизмы
Гипертрофия скелетных мышц (от греч. hyper — больше и trophe — питание) — это адаптационное увеличение объема или массы мышечной ткани в ответ на систематические физические нагрузки, которое происходит за счет увеличения поперечного сечения отдельных мышечных волокон без изменения их количества. Этот процесс представляет собой результат длительного превышения скорости синтеза мышечных белков над скоростью их распада, что приводит к накоплению белковой массы в мышечных клетках.
Гипертрофия является медленным процессом — измеримые изменения в объеме мышц становятся заметными только через 6-8 недель регулярных тренировок. Исследования показывают, что прирост мышечной массы у нетренированных людей может составлять 0,5-1,5 килограмма в месяц в первые три месяца тренировок, но затем эта скорость замедляется до 0,25-0,5 килограмма в месяц. У опытных атлетов годовой прирост мышечной массы редко превышает 2-3 килограмма чистой мышечной ткани.
Максимальный теоретический потенциал мышечной гипертрофии у человека ограничен генетическими факторами. У мужчин без использования фармакологии максимальный индекс мышечной массы (отношение мышечной массы к росту в квадрате) составляет около 25-27 кг/м, тогда как у женщин — 21-23 кг/м. Превышение этих значений возможно только при специфических генетических вариациях или применении анаболических стимуляторов.
В основе гипертрофии лежит фундаментальный биологический принцип суперкомпенсации — ответной реакции организма на повреждение или стресс. Физическая нагрузка создает три типа стресса для мышечных волокон: механический (натяжение и микроповреждения структур), метаболический (истощение энергетических запасов и накопление продуктов обмена) и гормональный (изменение концентрации анаболических и катаболических гормонов). Эти стрессоры запускают сложный каскад внутриклеточных сигнальных путей, которые в конечном итоге активируют синтез новых белков.
Существует несколько ключевых механизмов, запускающих гипертрофию. Механотрансдукция — процесс, при котором механическая нагрузка на мышечное волокно преобразуется в биохимические сигналы. Белки-сенсоры в мембране и цитоскелете мышечной клетки (интегрины, фокальные адгезионные комплексы, белки Z-дисков) реагируют на растяжение и деформацию, активируя внутриклеточные сигнальные пути. Основной из них — mTOR-путь (mechanistic target of rapamycin), который считается главным регулятором синтеза белка в скелетных мышцах.
Активация mTOR происходит в ответ на три основных стимула: механическую нагрузку на мышечные волокна, доступность аминокислот (особенно лейцина) и повышение уровня анаболических гормонов (инсулина, инсулиноподобного фактора роста-1). Когда mTOR активирован, он запускает трансляцию — процесс сборки новых белковых молекул на рибосомах. Одновременно mTOR подавляет аутофагию — процесс разрушения старых и поврежденных белков, что создает благоприятные условия для накопления белковой массы.
| Механизм гипертрофии | Тип стимула | Временная шкала активации | Вклад в прирост массы |
|---|---|---|---|
| Механическое напряжение | Высокая нагрузка (>65% от 1ПМ), растяжение под нагрузкой | Активация в течение минут, эффект до 48 часов | 40-50% |
| Метаболический стресс | Истощение креатинфосфата, накопление лактата и H+ | Активация в течение секунд, эффект до 24 часов | 25-35% |
| Микроповреждения | Эксцентрические нагрузки, новые движения | Развивается 6-48 часов, эффект до 7-10 дней | 15-25% |
| Гормональный ответ | Повышение тестостерона, гормона роста, IGF-1 | Пик через 15-60 минут, эффект до 24 часов | 10-20% (модулирующая роль) |
Критическую роль в гипертрофии играют клетки-сателлиты — миогенные стволовые клетки, которые располагаются между базальной мембраной и сарколеммой мышечных волокон. В покое эти клетки находятся в неактивном состоянии, но при повреждении или интенсивной стимуляции мышечного волокна они активируются, начинают делиться и сливаются с существующими волокнами, добавляя свои ядра. Это увеличивает количество миоядер в волокне, что расширяет его способность синтезировать белки — каждое ядро контролирует определенный объем цитоплазмы (миоядерный домен составляет около 2000 мкм).
Компромисс гипертрофии заключается в следующем: увеличение мышечной массы требует значительных энергетических затрат на синтез белков (около 4 молекул АТФ на одну пептидную связь) и повышенного потребления пищевого белка (минимум 1,6-2,2 грамма на килограмм массы тела в сутки). Кроме того, большие мышцы требуют больше кислорода и питательных веществ в состоянии покоя — базальный метаболизм увеличивается примерно на 50-100 килокалорий в сутки на каждый килограмм набранной мышечной массы. Для тех, кто начинает тренировочный путь, персональные тренировки в тренажерном зале помогают освоить правильную технику и построить эффективную программу для достижения гипертрофии.
Историческая эволюция понимания механизмов гипертрофии
До 1960-х годов считалось, что увеличение мышечной массы происходит за счет увеличения количества мышечных волокон (гиперплазия). Однако в 1970-е годы тщательные гистологические исследования на животных и людях показали, что количество волокон практически не меняется после рождения, а рост происходит за счет увеличения диаметра существующих волокон. В 1980-е годы были открыты клетки-сателлиты и их роль в регенерации мышц. В 1990-е годы начались молекулярные исследования сигнальных путей, а в 2000-е годы был детально изучен mTOR-путь, что революционизировало понимание механизмов роста мышц. Современные исследования 2010-2020-х годов показывают, что ограниченная гиперплазия возможна при экстремальных нагрузках (например, у элитных бодибилдеров), но ее вклад в общий прирост массы составляет менее 5%.
Существует три основные гипотезы, объясняющие запуск гипертрофии. Гипотеза механического повреждения предполагает, что микроразрывы сократительных белков и структур цитоскелета при интенсивных нагрузках запускают воспалительный ответ и последующую регенерацию с избыточным восстановлением. Исследования показывают, что даже однократная интенсивная тренировка может привести к повреждению более 80% мышечных волокон в тренируемой группе. Гипотеза метаболического стресса (ацидоза) указывает на то, что накопление лактата, ионов водорода и других метаболитов создает осмотический стресс, вызывающий набухание клеток и активацию анаболических сигнальных путей. Гипотеза гипоксии связывает гипертрофию с временным ограничением кровотока при напряжении мышцы более 60% от максимума, что вызывает накопление гипоксия-индуцируемого фактора-1α (HIF-1α) и активацию компенсаторных механизмов.
Современное понимание гипертрофии предполагает, что все три механизма работают синергетически, а их относительный вклад зависит от параметров тренировки. Тяжелые нагрузки (80-90% от максимума) с небольшим количеством повторений (3-6) создают преимущественно механический стресс. Умеренные нагрузки (65-75%) с средним диапазоном повторений (8-12) обеспечивают баланс механического и метаболического стресса. Легкие нагрузки (30-50%) с высоким количеством повторений (15-30) и ограничением кровотока создают выраженный метаболический стресс при меньшем механическом повреждении.
Миофибриллярная гипертрофия — рост за счет сократительных белков
Миофибриллярная гипертрофия — это увеличение объема мышечного волокна за счет роста количества и размера миофибрилл, содержащих сократительные белки актин и миозин, что приводит к прямому увеличению силы мышцы. При этом типе гипертрофии плотность миофибрилл в мышечном волокне возрастает, а относительный объем саркоплазмы (несократительной части клетки) может даже уменьшаться или оставаться постоянным.
Миофибриллярная гипертрофия представляет собой адаптацию к высокоинтенсивным силовым нагрузкам с большими весами и относительно малым количеством повторений. Главным стимулом для этого типа роста является высокое механическое напряжение — произведение силы сокращения на время под нагрузкой. Когда мышечное волокно генерирует силу близкую к максимальной (выше 70-80% от одноповторного максимума), активируются механосенсоры в Z-дисках миофибрилл, которые преобразуют механический сигнал в биохимический каскад.
Ключевые белки, участвующие в механотрансдукции при миофибриллярной гипертрофии, включают тайтин (гигантский белок, соединяющий Z-диск с М-линией и работающий как молекулярная пружина), небулин (белок, определяющий длину тонких филаментов) и комплекс белков костамеров, которые связывают Z-диски с сарколеммой. При высоком механическом напряжении эти белки деформируются, что открывает скрытые сайты связывания для сигнальных молекул и активирует каскад фосфорилирования через киназы семейства MAPK (митоген-активируемые протеинкиназы).
Исследования биопсий мышц пауэрлифтеров и тяжелоатлетов показывают, что у этих атлетов плотность миофибрилл в мышечных волокнах типа II на 15-25% выше, чем у нетренированных людей, а площадь поперечного сечения отдельных миофибрилл увеличена на 20-30%. При этом объем саркоплазмы на единицу объема волокна у силовых атлетов на 10-15% ниже, чем у бодибилдеров.
Процесс сборки новых миофибрилл происходит по принципу строительных лесов. Сначала в саркоплазме образуются небольшие структуры, называемые премиофибриллы или стресс-фибриллы, диаметром 0,1-0,2 микрометра. Они содержат неполностью организованные актиновые и миозиновые филаменты в виде небольших регистров. Эти структуры постепенно встраиваются между существующими миофибриллами и созревают, увеличиваясь в диаметре до 1-2 микрометров — размера зрелой миофибриллы. Весь процесс от начала синтеза белков до формирования полноценной сократительной единицы занимает 7-14 дней.
Синтез сократительных белков регулируется на нескольких уровнях. На уровне транскрипции происходит считывание генов, кодирующих миозин тяжелых цепей (MHC), актин, тропонин и тропомиозин. Этот процесс активируется транскрипционными факторами семейства MEF2 (myocyte enhancer factor 2) и NFAT (nuclear factor of activated T-cells), которые транслоцируются в ядро в ответ на повышение концентрации кальция и активацию кальцинейрина. На уровне трансляции работает mTOR-комплекс 1, который фосфорилирует рибосомальный белок S6K1 и фактор инициации трансляции 4E-BP1, ускоряя сборку белков на рибосомах в 2-3 раза.
| Компонент миофибриллы | Основные белки | Функция | Скорость обновления |
|---|---|---|---|
| Толстые филаменты | Миозин (тяжелые и легкие цепи) | Генерация силы через гидролиз АТФ | 30-45 дней (период полураспада) |
| Тонкие филаменты | Актин, тропонин, тропомиозин | Взаимодействие с миозином, регуляция Ca2+ | 7-14 дней |
| Z-диски | α-актинин, капа-белки, филамин | Закрепление тонких филаментов, передача силы | 14-21 день |
| М-линия | Миомезин, M-протеин | Стабилизация толстых филаментов | 30-60 дней |
| Эластичные элементы | Тайтин, небулин | Пассивная жесткость, регуляция длины | 45-90 дней |
Важным аспектом миофибриллярной гипертрофии является изменение изоформного состава миозина. В скелетных мышцах человека экспрессируются три основные изоформы тяжелых цепей миозина: MHC-I (медленный окислительный тип), MHC-IIA (быстрый окислительно-гликолитический) и MHC-IIX (быстрый гликолитический). Силовые тренировки с тяжелыми весами индуцируют переход волокон от MHC-IIX к MHC-IIA — более медленной, но более выносливой изоформе. Это может показаться парадоксальным, но на самом деле представляет собой оптимизацию: волокна типа IIA генерируют почти такую же максимальную силу, как IIX (разница менее 10%), но при этом обладают в 2-3 раза большей устойчивостью к утомлению благодаря лучшему митохондриальному оснащению.
Прирост силы при миофибриллярной гипертрофии происходит по нескольким причинам. Во-первых, увеличение количества миофибрилл прямо увеличивает число поперечных мостиков, которые могут взаимодействовать с актином одновременно — каждая дополнительная миофибрилла добавляет сотни тысяч потенциальных точек генерации силы. Во-вторых, увеличивается жесткость сухожилий — адаптация, которая позволяет более эффективно передавать генерируемую мышцей силу на кости. В-третьих, происходит улучшение нервно-мышечной координации — повышается частота импульсации мотонейронов и синхронизация активации двигательных единиц.
Количественная связь между площадью поперечного сечения и силой
Теоретически сила мышцы должна быть прямо пропорциональна площади ее физиологического поперечного сечения (PCSA — physiological cross-sectional area). Экспериментальные данные показывают, что скелетные мышцы человека генерируют около 25-35 ньютонов силы на квадратный сантиметр PCSA. Однако связь между гипертрофией и силой не всегда линейна. У начинающих атлетов в первые 6-8 недель тренировок прирост силы на 40-60% происходит за счет нейронных адаптаций (улучшение рекрутирования двигательных единиц) при минимальной гипертрофии. У продвинутых атлетов, наоборот, дальнейший прирост силы почти полностью зависит от гипертрофии — каждый 1% прироста мышечной массы дает примерно 0,5-0,7% прироста максимальной силы. Расхождение объясняется тем, что у опытных спортсменов нейронные факторы уже оптимизированы, и дальнейший прогресс лимитирован исключительно количеством сократительного материала.
Для максимальной стимуляции миофибриллярной гипертрофии оптимальными считаются нагрузки 75-90% от одноповторного максимума с 3-6 повторениями в подходе, отдыхом 2-5 минут между подходами и акцентом на многосуставные базовые упражнения (приседания, становая тяга, жим лежа). Эксцентрическая фаза движения (опускание веса) должна выполняться под контролем в течение 2-4 секунд, что создает высокое механическое напряжение при удлинении мышцы. Частота тренировок для одной мышечной группы — 2-3 раза в неделю с интервалом не менее 48 часов для полного восстановления синтеза белка. Построить такую программу помогут специалисты, которые работают в силовых программах тренировок.
Компромисс миофибриллярной гипертрофии заключается в том, что максимальный прирост сократительных белков требует высоких нагрузок, которые создают значительный стресс для суставов, связок и центральной нервной системы. Тренировки с весами 85-95% от максимума могут выполняться только 2-3 раза в неделю из-за длительного (5-7 дней) восстановления нейромышечной системы. Кроме того, быстрая миофибриллярная гипертрофия без соответствующего увеличения капилляризации может привести к относительному снижению плотности капилляров на единицу мышечной массы, что ухудшает доставку кислорода и питательных веществ при длительной работе.

Саркоплазматическая гипертрофия — увеличение объема несократительных структур
Саркоплазматическая гипертрофия — это увеличение объема мышечного волокна за счет роста несократительных компонентов саркоплазмы: запасов гликогена и креатинфосфата, объема саркоплазматического ретикулума, митохондрий, миоглобина и других белков метаболизма, при этом количество миофибрилл увеличивается в меньшей степени или остается неизменным. Этот тип гипертрофии приводит к увеличению выносливости мышцы и ее способности выполнять повторную работу, но дает меньший прирост максимальной силы на единицу массы.
Главным стимулом для саркоплазматической гипертрофии является метаболический стресс — истощение энергетических субстратов и накопление метаболитов при длительной или высокообъемной работе. Когда мышца выполняет серию сокращений с умеренной интенсивностью (60-75% от максимума) и короткими периодами отдыха (30-90 секунд), запасы креатинфосфата истощаются за 10-15 секунд, концентрация АТФ падает на 30-50%, а концентрация неорганического фосфата и АДФ возрастает в 3-5 раз. Гликоген расщепляется анаэробным путем, что приводит к накоплению лактата и снижению pH с 7,0 до 6,5-6,3.
Эти метаболические изменения запускают специфические сигнальные пути. Снижение соотношения АТФ/АМФ активирует AMPK (AMP-activated protein kinase) — энергетический сенсор клетки, который стимулирует процессы, направленные на производство энергии. AMPK увеличивает экспрессию генов митохондриальных белков через коактиватор PGC-1α (peroxisome proliferator-activated receptor gamma coactivator 1-alpha), что приводит к росту числа и размера митохондрий — процессу, называемому митохондриальный биогенез. Одновременно AMPK стимулирует синтез белков, участвующих в транспорте глюкозы (GLUT4) и запасании гликогена (гликоген-синтаза).
У бодибилдеров, тренирующихся в высокообъемном режиме с умеренными весами, концентрация гликогена в мышцах может достигать 40-50 граммов на килограмм мышечной ткани (в 2-2,5 раза выше, чем у нетренированных людей). Поскольку каждый грамм гликогена связывает 3-4 грамма воды, это дает прирост объема мышц на 15-20% без увеличения количества сократительных белков.
Накопление метаболитов, особенно ионов водорода и неорганического фосфата, создает осмотический стресс — повышение концентрации растворенных веществ в саркоплазме. Это приводит к притоку воды в мышечную клетку и ее набуханию. Набухание клетки само по себе является анаболическим сигналом — оно активирует интегрины в мембране и сигнальные пути, стимулирующие синтез белка и подавляющие его распад. Этот механизм можно сравнить с надуванием воздушного шара — увеличение внутреннего объема создает механическое напряжение на оболочку, что воспринимается клеткой как стимул для укрепления структур.
Важным компонентом саркоплазматической гипертрофии является увеличение объема саркоплазматического ретикулума. При тренировках с высоким числом повторений (12-20) мышца должна многократно сокращаться и расслабляться, что требует эффективной работы SERCA-насосов для быстрого удаления кальция из саркоплазмы. В ответ на этот стимул клетка синтезирует дополнительные мембраны СР и увеличивает экспрессию SERCA. Исследования показывают, что у спортсменов на выносливость плотность SERCA в мышцах на 30-40% выше, а общий объем СР на 20-25% больше по сравнению с силовыми атлетами.
- Миоглобин
- Кислородосвязывающий белок в саркоплазме, который увеличивает запас кислорода в мышце и ускоряет его диффузию к митохондриям. При саркоплазматической гипертрофии концентрация миоглобина может возрастать на 50-80%.
- Капилляризация
- Увеличение количества капилляров на единицу площади поперечного сечения мышцы. При метаболическом стрессе высвобождается фактор роста эндотелия сосудов (VEGF), стимулирующий образование новых капилляров — плотность капилляров может увеличиться с 300-400 до 500-600 на мм.
- Буферная емкость
- Способность мышцы нейтрализовать ионы водорода. Увеличивается концентрация буферных систем (карнозин, фосфатная буферная система), что позволяет дольше поддерживать высокую интенсивность работы при накоплении лактата.
Саркоплазматическая гипертрофия происходит преимущественно в волокнах типа I (медленных окислительных) и типа IIA (быстрых окислительно-гликолитических), которые обладают большей капилляризацией и метаболической пластичностью. Волокна типа IIX (быстрые гликолитические) также участвуют в этой адаптации, но в меньшей степени. Интересно, что длительные тренировки в высокообъемном режиме могут индуцировать переход волокон IIX → IIA → I, что увеличивает долю медленных волокон и улучшает окислительный потенциал мышцы.
| Компонент саркоплазмы | Исходное содержание | После адаптации | Функциональное значение |
|---|---|---|---|
| Гликоген | 15-20 г/кг мышцы | 40-50 г/кг мышцы | Энергетический субстрат для анаэробной и аэробной работы |
| Креатинфосфат | 17-20 ммоль/кг | 25-30 ммоль/кг | Быстрое ресинтезирование АТФ в первые 10-15 секунд работы |
| Митохондрии | 3-5% объема волокна | 6-10% объема волокна | Окислительное фосфорилирование, выносливость |
| Саркоплазматический ретикулум | 5-7% объема волокна | 7-10% объема волокна | Управление кальцием, скорость сокращения/расслабления |
| Миоглобин | 5-8 мг/г ткани | 10-15 мг/г ткани | Резерв кислорода, диффузия O2 к митохондриям |
Оптимальные параметры тренировки для стимуляции саркоплазматической гипертрофии включают умеренную интенсивность (60-75% от максимума), высокое число повторений (12-20 и более), короткие периоды отдыха между подходами (30-90 секунд) и высокий общий объем работы (15-25 рабочих подходов на мышечную группу в неделю). Техника выполнения должна обеспечивать постоянное мышечное напряжение без полного расслабления в верхней и нижней точках движения — метод непрерывного напряжения. Дополнительные методы включают дропсеты, суперсеты и тренировки с ограничением кровотока (BFR — blood flow restriction).
Феномен "памп-эффекта" и его связь с гипертрофией
Памп (от англ. pump — накачка) — временное увеличение объема мышцы во время и сразу после тренировки, связанное с притоком крови и задержкой жидкости в мышечной ткани. Раньше считалось, что памп — это чисто косметический эффект без анаболического значения. Однако современные исследования показывают, что клеточное набухание является мощным анаболическим сигналом, активирующим синтез белка и подавляющим его распад через интегрин-связанные сигнальные пути. Хронический памп, возникающий при регулярных высокообъемных тренировках, приводит к долговременному увеличению запасов гликогена и связанной с ним воды, что вносит значительный вклад в саркоплазматическую гипертрофию. У профессиональных бодибилдеров до 30-40% объема мышц может приходиться на несократительные компоненты.
Компромисс саркоплазматической гипертрофии заключается в том, что увеличение объема мышц не сопровождается пропорциональным приростом максимальной силы. Плотность миофибрилл (количество сократительного материала на единицу объема) при этом типе гипертрофии снижается на 10-20%. Это означает, что атлет с выраженной саркоплазматической гипертрофией может иметь впечатляющий мышечный объем, но относительно низкие силовые показатели на килограмм массы тела по сравнению с пауэрлифтером аналогичного веса. С другой стороны, повышенная капилляризация, митохондриальная плотность и запасы гликогена обеспечивают лучшую работоспособность в высокообъемных тренировках и более быстрое восстановление между подходами.
Существование саркоплазматической гипертрофии как отдельного явления долгое время было предметом дискуссии. Некоторые исследователи утверждали, что наблюдаемые различия между бодибилдерами и силовыми атлетами объясняются временным отеком и задержкой жидкости, а не стабильными структурными изменениями. Однако биопсийные исследования 2010-2020-х годов убедительно показали, что у атлетов, тренирующихся в высокообъемном режиме, действительно наблюдается долговременное увеличение несократительных компонентов клетки — повышенное содержание гликогена сохраняется даже через 48-72 часа после последней тренировки, а плотность митохондрий и объем СР остаются повышенными в течение недель после прекращения тренировок.
Как различаются миофибриллярная и саркоплазматическая гипертрофия по функциональным результатам
Миофибриллярная и саркоплазматическая гипертрофия приводят к принципиально разным функциональным адаптациям: первая увеличивает максимальную силу и мощность за счет роста сократительных белков, вторая повышает работоспособность и выносливость за счет улучшения энергетического обеспечения, при этом прирост мышечной массы на один килограмм при миофибриллярной гипертрофии дает в 1,5-2 раза больший прирост силы, чем при саркоплазматической.
Ключевое различие между двумя типами гипертрофии заключается в изменении состава мышечного волокна. При миофибриллярной гипертрофии доля сократительных белков (актина и миозина) возрастает с 55-60% до 65-70% от общего объема волокна, что означает увеличение плотности упаковки миофибрилл. У элитных пауэрлифтеров количество миофибрилл на квадратный миллиметр поперечного сечения мышцы может достигать 800-1000, тогда как у нетренированных людей этот показатель составляет 400-600. При саркоплазматической гипертрофии, наоборот, относительный объем саркоплазмы увеличивается с 35-40% до 45-50%, а доля сократительных белков снижается.
Функциональная разница проявляется в показателях удельной силы — силы, генерируемой на единицу площади поперечного сечения мышцы. Исследования 2015-2020 годов показали, что пауэрлифтеры и тяжелоатлеты, у которых доминирует миофибриллярная гипертрофия, демонстрируют удельную силу 35-40 ньютонов на квадратный сантиметр, тогда как у бодибилдеров с выраженной саркоплазматической гипертрофией этот показатель составляет 25-30 Н/см. Разница в 25-40% означает, что при одинаковом размере мышц силовой атлет будет значительно сильнее.
Сравнительное исследование биопсий мышц у 40 элитных атлетов разных специализаций (пауэрлифтинг, бодибилдинг, кроссфит) показало, что у пауэрлифтеров площадь поперечного сечения отдельных миофибрилл была на 23% больше, а их плотность — на 18% выше по сравнению с бодибилдерами при идентичном общем размере мышечных волокон. При этом у бодибилдеров содержание гликогена было выше на 45%, а плотность капилляров — на 35%.
Различия в энергетическом метаболизме между двумя типами гипертрофии также значительны. При саркоплазматической гипертрофии увеличивается буферная емкость мышцы — способность нейтрализовать ионы водорода и поддерживать pH близким к нейтральному. Концентрация карнозина (дипептид, работающий как внутриклеточный буфер) может возрастать с 15-20 до 30-35 ммоль на килограмм сухой массы. Это позволяет мышце дольше работать в условиях накопления лактата — критический фактор для высокообъемных тренировок с короткими периодами отдыха.
| Параметр | Миофибриллярная гипертрофия | Саркоплазматическая гипертрофия | Функциональное значение |
|---|---|---|---|
| Удельная сила (Н/см) | 35-40 | 25-30 | Способность генерировать максимальную силу |
| Плотность миофибрилл (на мм) | 800-1000 | 600-750 | Количество сократительного материала |
| Содержание гликогена (г/кг) | 20-30 | 40-50 | Запас энергии для длительной работы |
| Плотность капилляров (на мм) | 350-450 | 500-650 | Доставка кислорода и питательных веществ |
| Объем митохондрий (% от волокна) | 3-5% | 7-12% | Аэробная производительность |
| Время до утомления (при 70% max) | 20-30 секунд | 45-90 секунд | Способность к повторной работе |
Нейромышечные адаптации также различаются между типами гипертрофии. Миофибриллярная гипертрофия сопровождается повышением частоты импульсации мотонейронов — нервные клетки, иннервирующие мышцы, учатся посылать импульсы с большей частотой (до 80-120 Гц вместо базовых 30-50 Гц). Это позволяет рекрутировать больше двигательных единиц одновременно и синхронизировать их работу. При саркоплазматической гипертрофии нейронные адаптации выражены слабее, но улучшается нейромышечная выносливость — способность поддерживать субмаксимальные сокращения длительное время без снижения частоты импульсации.
Интересный феномен — различия в скорости сокращения мышц при разных типах гипертрофии. Миофибриллярная гипертрофия, особенно при тренировках с максимальными весами, приводит к небольшому увеличению времени достижения пикового напряжения (на 5-10%), поскольку большим мышечным волокнам требуется больше времени для активации всех миофибрилл. Саркоплазматическая гипертрофия, наоборот, может ускорять расслабление мышцы благодаря увеличению объема саркоплазматического ретикулума и плотности SERCA-насосов — время полурасслабления может сокращаться с 50-60 миллисекунд до 35-45 миллисекунд.
Можно ли изменить тип гипертрофии переключением тренировочного режима
Тип гипертрофии не является фиксированным — он может изменяться в зависимости от тренировочного стимула. Если атлет с выраженной миофибриллярной гипертрофией (например, пауэрлифтер) переключается на высокообъемный режим тренировок с умеренными весами, в течение 8-12 недель происходит значительное увеличение несократительных компонентов — запасы гликогена возрастают на 40-60%, плотность капилляров увеличивается на 25-35%. При этом количество миофибрилл остается практически неизменным. Обратный переход (от саркоплазматической к миофибриллярной) также возможен, но происходит медленнее — требуется 12-16 недель тяжелых силовых тренировок для значимого прироста плотности миофибрилл. Это объясняется тем, что синтез сократительных белков — более длительный процесс (период полураспада миозина составляет 30-45 дней), чем накопление гликогена или рост митохондрий.
Практическое значение различий между типами гипертрофии велико для построения тренировочной программы. Атлеты, цель которых — максимальная сила (пауэрлифтинг, тяжелая атлетика, единоборства), должны приоритизировать миофибриллярную гипертрофию, используя тяжелые веса (80-95% от максимума), малое количество повторений (1-5) и длительные периоды отдыха (3-5 минут). Те, кто стремится к максимальному мышечному объему с хорошей работоспособностью (бодибилдинг, фитнес), получат больше пользы от комбинированного подхода с акцентом на саркоплазматическую гипертрофию — умеренные веса (65-75%), высокое число повторений (10-20) и короткие периоды отдыха (60-90 секунд).
Компромисс выбора между типами гипертрофии состоит в том, что невозможно одновременно максимизировать оба направления адаптации. Тренировки, оптимальные для миофибриллярной гипертрофии, создают недостаточный метаболический стресс для значительного увеличения несократительных компонентов. Тренировки, стимулирующие саркоплазматическую гипертрофию, недостаточно интенсивны для максимального прироста сократительных белков. Наиболее эффективная стратегия для большинства атлетов — периодизация, когда фазы акцента на силу (4-6 недель тяжелых тренировок) чередуются с фазами акцента на объем (4-6 недель высокообъемных тренировок), что позволяет развивать оба типа адаптации последовательно.

Влияние на силовые показатели: почему миофибриллы важнее объема
Максимальная сила мышцы определяется в первую очередь количеством и качеством миофибрилл, а не общим объемом мышечного волокна, поскольку только сократительные белки актин и миозин непосредственно генерируют силу через образование поперечных мостиков и гидролиз АТФ. Увеличение объема мышцы за счет несократительных компонентов (гликоген, вода, митохондрии) практически не влияет на максимальную силу — корреляция между размером мышцы и силой составляет r=0,6-0,7, что означает, что размер объясняет только 35-50% вариабельности силовых показателей.
Физиологическая основа этого явления кроется в механизме мышечного сокращения. Сила, которую может генерировать мышечное волокно, прямо пропорциональна числу одновременно работающих поперечных мостиков между актином и миозином. Каждый мостик при оптимальной длине саркомера генерирует силу около 3-5 пиконьютонов. В одной зрелой миофибрилле диаметром 1-2 микрометра содержится примерно 1000-1500 молекул миозина, что дает около 2000-3000 потенциальных точек прикрепления. Увеличение количества миофибрилл на 20% прямо увеличивает максимальную силу на 20%, тогда как увеличение объема саркоплазмы на 20% не добавляет ни одного поперечного мостика.
Классическое исследование Икегавы и коллег (2008) на японских тяжелоатлетах показало, что при одинаковой окружности бедра (60-62 см) максимальная сила в приседании варьировалась от 180 до 260 кг — разница в 44%. Биопсия четырехглавой мышцы выявила, что у более сильных атлетов плотность миофибрилл была на 28% выше, а содержание гликогена и объем митохондрий не различались значимо между группами.
Существует несколько механизмов, по которым миофибриллярная плотность влияет на силовые показатели. Первый — геометрический эффект: при большем количестве миофибрилл увеличивается площадь физиологического поперечного сечения (PCSA), что является главным детерминантом максимальной силы. Исследования на изолированных мышечных волокнах показывают линейную зависимость: каждый квадратный сантиметр PCSA генерирует 25-35 ньютонов силы при оптимальных условиях (температура 37°C, оптимальная длина саркомеров 2,4-2,6 микрометров, полная активация). Второй механизм — оптимизация передачи силы: плотно упакованные миофибриллы создают более жесткую внутреннюю структуру волокна, что уменьшает потери силы на деформацию упругих элементов.
Нейронные факторы усиливают эффект миофибриллярной плотности на силу. При тренировках с максимальными весами (90-100% от одноповторного максимума) происходит адаптация центральной нервной системы, включающая увеличение моторной единицы рекрутмента — способности активировать высокопороговые быстрые двигательные единицы, содержащие наибольшее количество миофибрилл. У нетренированных людей произвольная активация мышцы достигает только 70-80% от максимально возможной (определяемой электростимуляцией), тогда как у элитных силовых атлетов этот показатель может достигать 95-98%. Однако эта нейронная адаптация эффективна только при наличии достаточного количества сократительного материала.
| Фактор силы | Вклад в максимальную силу | Зависимость от миофибрилл | Время развития адаптации |
|---|---|---|---|
| Площадь поперечного сечения миофибрилл | 40-50% | Прямая зависимость | 8-12 недель |
| Нейронная активация (рекрутмент) | 20-30% | Косвенная (реализует потенциал миофибрилл) | 4-8 недель |
| Жесткость сухожилий | 10-15% | Независима от миофибрилл | 12-24 недели |
| Архитектура мышцы (угол перистости) | 10-15% | Изменяется при росте миофибрилл | 8-16 недель |
| Тип миозиновых изоформ | 5-10% | Качество миофибрилл | 6-12 недель |
| Энергетическое обеспечение | <5% | Не зависит от количества миофибрилл | 2-4 недели |
Качество миофибрилл также влияет на силу не меньше, чем их количество. Изоформный состав миозина определяет скорость и силу сокращения: волокна с миозином типа IIX генерируют на 15-20% большую силу и в 3-4 раза большую мощность, чем волокна с миозином типа I при одинаковом размере. Силовые тренировки индуцируют гипертрофию преимущественно быстрых волокон типа II — у элитных пауэрлифтеров до 60-70% волокон в основных работающих мышцах (квадрицепс, большая ягодичная) принадлежат к типу II, тогда как у нетренированных людей это соотношение составляет 45-55%.
Практический пример демонстрирует важность миофибриллярной плотности для силы. Два атлета имеют одинаковую окружность руки (40 см) и одинаковую массу тела (90 кг). Атлет А — пауэрлифтер, который тренируется с весами 85-95% от максимума, выполняя 3-5 повторений с отдыхом 3-5 минут. Атлет Б — бодибилдер, тренирующийся с весами 65-75% от максимума, выполняя 10-15 повторений с отдыхом 60-90 секунд. В жиме лежа атлет А поднимает 150 кг на одно повторение, атлет Б — 110 кг. Разница в 36% объясняется тем, что у атлета А плотность миофибрилл на 25-30% выше, нейронная активация лучше на 10-15%, а жесткость сухожилий больше на 15-20%.
Почему бодибилдеры слабее пауэрлифтеров при большем размере мышц
Это кажущийся парадокс имеет простое объяснение. Профессиональные бодибилдеры действительно имеют большие мышцы, но значительная часть их объема приходится на несократительные компоненты: гликоген (40-50 г/кг вместо 20-30 г/кг), связанная с гликогеном вода (в 3-4 раза больше массы гликогена), увеличенный объем саркоплазматического ретикулума и митохондрий. У элитного бодибилдера с окружностью руки 50 см фактическая площадь поперечного сечения миофибрилл может быть такой же или даже меньше, чем у пауэрлифтера с окружностью руки 45 см. Кроме того, бодибилдеры не тренируют максимальную нейронную активацию — их тренировки направлены на метаболический стресс, а не на рекрутмент высокопороговых двигательных единиц. Результат: при тестировании максимальной силы бодибилдер весом 100 кг может уступать пауэрлифтеру весом 90 кг на 20-30% в базовых упражнениях, несмотря на визуально большие мышцы.
Существует верхний предел, до которого увеличение миофибриллярной плотности может повышать силу. При очень высокой плотности упаковки (более 900-1000 миофибрилл на квадратный миллиметр) начинаются проблемы с доставкой кислорода и питательных веществ к центральным миофибриллам из-за увеличения диффузионного расстояния. Это один из факторов, ограничивающих максимальный размер мышечных волокон — волокна диаметром более 120-150 микрометров испытывают гипоксию центральных областей при интенсивной работе. Эволюционное решение этой проблемы — не бесконечное увеличение размера отдельных волокон, а рост числа волокон через гиперплазию, но у человека этот механизм работает крайне ограниченно.
Для максимизации силовых показателей программа тренировок должна приоритизировать стимулы, специфичные для миофибриллярной гипертрофии. Оптимальные параметры включают интенсивность 80-95% от одноповторного максимума, объем 3-6 повторений в подходе, 3-5 подходов на упражнение, отдых 3-5 минут между подходами для полного восстановления креатинфосфата. Частота тренировок — 2-3 раза в неделю на мышечную группу. Акцент на многосуставные базовые движения (приседания, становая тяга, жим лежа, подтягивания), которые позволяют использовать максимальные веса и создают высокое механическое напряжение. Профессиональное руководство помогут получить индивидуальные тренировки с тренером, где программа адаптируется под конкретные силовые цели.
Цена специализации на максимальной силе через миофибриллярную гипертрофию включает повышенный риск травм соединительной ткани (сухожилий, связок) из-за высоких нагрузок, длительное восстановление центральной нервной системы (5-7 дней после тренировки с весами выше 90%), относительно низкую работоспособность в высокообъемных режимах из-за недостаточного развития энергетических систем. Атлеты, ориентированные исключительно на максимальную силу, часто имеют сниженную капилляризацию мышц (на 20-30% ниже, чем у спортсменов на выносливость) и меньшую плотность митохондрий, что ограничивает их способность к длительной работе субмаксимальной интенсивности.
Влияние на выносливость: роль запасов гликогена и митохондрий
Выносливость мышцы определяется в первую очередь энергетическим обеспечением — способностью поддерживать ресинтез АТФ в течение длительного времени, что зависит от запасов гликогена, плотности митохондрий и эффективности доставки кислорода через капиллярную сеть. Саркоплазматическая гипертрофия, увеличивающая объем несократительных компонентов, может повышать мышечную выносливость на 40-80% без значительного роста максимальной силы, что делает ее критически важной для видов спорта, требующих повторных усилий высокой интенсивности.
Гликоген является главным энергетическим субстратом для работы мышц интенсивностью выше 60-70% от максимума. В покоящейся мышце нетренированного человека содержится 15-20 граммов гликогена на килограмм мышечной ткани, что эквивалентно примерно 300-400 ммоль глюкозных единиц на килограмм. При высокообъемных тренировках с умеренными весами и короткими периодами отдыха мышца адаптируется, увеличивая запасы гликогена до 40-50 г/кг — прирост в 2-2,5 раза. Эта адаптация происходит через повышение экспрессии гликоген-синтазы и увеличение числа молекул переносчика глюкозы GLUT4 в мембране.
Физиологическое значение увеличенных запасов гликогена многогранно. Во-первых, гликоген обеспечивает быстрый ресинтез АТФ через анаэробный гликолиз со скоростью до 2,5-3,0 ммоль АТФ на килограмм в секунду — в 2-3 раза быстрее, чем окислительное фосфорилирование. Это критично для работы интенсивностью 70-90% от максимума, которая не может полностью обеспечиваться аэробными процессами. Во-вторых, локальные запасы гликогена непосредственно в работающих мышцах позволяют избежать зависимости от доставки глюкозы через кровь, что становится лимитирующим фактором при длительной работе.
Исследование Бергстрома и Хультмана (1967) — классическая работа по физиологии мышц — показало, что истощение гликогена в мышце коррелирует с развитием утомления сильнее, чем любой другой метаболический параметр. Когда концентрация гликогена падает ниже 50 ммоль/кг (примерно 8-10 г/кг), интенсивность работы снижается на 40-60% независимо от волевых усилий. Атлеты с высокими исходными запасами гликогена (40-50 г/кг) могут поддерживать высокоинтенсивную работу в 2-3 раза дольше, чем атлеты с низкими запасами (15-20 г/кг).
Каждый грамм гликогена связывает 3-4 грамма воды осмотическим эффектом, что имеет двойное функциональное значение. С одной стороны, эта связанная вода увеличивает объем мышцы на 10-20% и создает эффект наполненности — мышцы выглядят больше и плотнее. С другой стороны, эта вода служит резервуаром для терморегуляции и поддержания объема плазмы крови при интенсивной работе. При длительных тренировках в жару атлеты с высокими запасами гликогена и связанной воды демонстрируют на 15-25% лучшую работоспособность благодаря меньшему обезвоживанию.
Митохондрии представляют собой второй критический компонент выносливости. Эти органеллы выполняют окислительное фосфорилирование — процесс, в котором энергия окисления углеводов и жиров преобразуется в АТФ с эффективностью около 38-40%. В нетренированных мышечных волокнах типа II митохондрии занимают 3-5% объема, тогда как при саркоплазматической гипертрофии этот показатель может возрастать до 8-12% — увеличение в 2-3 раза. Это достигается через активацию транскрипционного коактиватора PGC-1α, который запускает экспрессию сотен генов митохондриальных белков.
| Компонент энергообеспечения | Нетренированная мышца | После саркоплазматической гипертрофии | Функциональный эффект |
|---|---|---|---|
| Гликоген (г/кг мышцы) | 15-20 | 40-50 | Длительность работы высокой интенсивности увеличена в 2-2,5 раза |
| Креатинфосфат (ммоль/кг) | 17-20 | 25-30 | Быстрое восстановление между повторениями (10-15% быстрее) |
| Объем митохондрий (% от волокна) | 3-5% | 8-12% | Аэробная мощность на 60-100% выше |
| Плотность капилляров (на мм) | 300-400 | 500-650 | Доставка O₂ и удаление метаболитов на 40-60% эффективнее |
| Активность окислительных ферментов | Базовая (100%) | 180-250% | Скорость аэробного ресинтеза АТФ удвоена |
| Буферная емкость (ммоль H⁺/кг) | 120-150 | 180-220 | Устойчивость к закислению на 30-50% выше |
Увеличение митохондриальной плотности имеет множественные функциональные преимущества. Во-первых, повышается аэробная мощность — максимальная скорость потребления кислорода мышцей, что позволяет работать с большей интенсивностью при использовании преимущественно аэробного метаболизма. Во-вторых, улучшается экономичность — для выполнения той же работы требуется меньше энергии и кислорода благодаря более эффективному сопряжению окисления и фосфорилирования. В-третьих, ускоряется восстановление между повторениями и подходами за счет более быстрого ресинтеза креатинфосфата, который происходит аэробным путем.
Капилляризация мышц является третьим компонентом, определяющим выносливость. При метаболическом стрессе, характерном для высокообъемных тренировок, мышечные волокна выделяют фактор роста эндотелия сосудов (VEGF) и другие ангиогенные факторы, стимулирующие образование новых капилляров. Плотность капилляров может увеличиться с 300-400 до 500-650 на квадратный миллиметр поперечного сечения — прирост на 50-80%. Это сокращает среднее диффузионное расстояние от капилляра до центра мышечного волокна с 25-30 до 15-20 микрометров, что радикально улучшает доставку кислорода и питательных веществ.
Практический пример демонстрирует важность энергетических адаптаций для выносливости. Атлет выполняет подходы приседаний с весом 70% от максимума (150 кг при максимуме 215 кг) с 60-секундным отдыхом между подходами. До начала тренировочной программы, направленной на саркоплазматическую гипертрофию, атлет мог выполнить 4 подхода по 10 повторений (всего 40 повторений) прежде чем наступало выраженное утомление. После 12 недель высокообъемных тренировок запасы гликогена увеличились на 60%, митохондриальная плотность — на 45%, капилляризация — на 35%. Результат: атлет может выполнить 7 подходов по 12 повторений (84 повторения) — увеличение работоспособности на 110% при неизменной максимальной силе.
Роль жирового метаболизма в мышечной выносливости
При работе интенсивностью ниже 65-70% от максимума значительную роль начинает играть окисление жирных кислот как источник энергии. Саркоплазматическая гипертрофия повышает способность мышцы использовать жиры через увеличение плотности митохондрий и активности ферментов β-окисления. У тренированных атлетов окисление жиров может обеспечивать до 60-70% энергетических потребностей при умеренной интенсивности работы (50-65% от максимума), тогда как у нетренированных — только 30-40%. Это гликоген-сберегающий эффект — способность экономить гликоген для критических моментов высокой интенсивности. Внутримышечные триглицериды (липидные капли в саркоплазме) также увеличиваются при тренировках на выносливость — с 5-10 до 15-25 ммоль/кг, что обеспечивает дополнительный локальный источник энергии. Однако при интенсивности выше 75% от максимума скорость окисления жиров падает из-за недостатка кислорода, и гликоген становится доминирующим субстратом.
Тренировки, оптимальные для развития энергетических адаптаций и мышечной выносливости, существенно отличаются от силовых протоколов. Оптимальные параметры включают умеренную интенсивность (60-75% от максимума), высокое число повторений (12-25), короткие периоды отдыха (30-90 секунд), высокий объем (15-25 рабочих подходов на мышечную группу в неделю). Методы, усиливающие метаболический стресс, особенно эффективны: суперсеты (два упражнения подряд без отдыха), дропсеты (снижение веса после отказа и продолжение), тренировки с ограничением кровотока (использование жгутов для создания гипоксии при 30-50% от максимума). Для восстановления энергетических запасов после таких тренировок критично адекватное потребление углеводов — 5-7 граммов на килограмм массы тела в сутки, а также использование энергетических батончиков для быстрого восполнения гликогена.
Компромисс специализации на выносливости через саркоплазматическую гипертрофию заключается в том, что увеличение объема несократительных компонентов снижает относительную плотность миофибрилл и, соответственно, удельную силу мышцы. Атлет с выраженной саркоплазматической гипертрофией будет иметь на 20-30% более низкие показатели максимальной силы на килограмм мышечной массы по сравнению со специализирующимся на миофибриллярной гипертрофии. Кроме того, большие запасы гликогена и связанной воды увеличивают массу тела на 2-4 кг, что может быть нежелательно в видах спорта с весовыми категориями или где важна относительная сила (гимнастика, скалолазание).
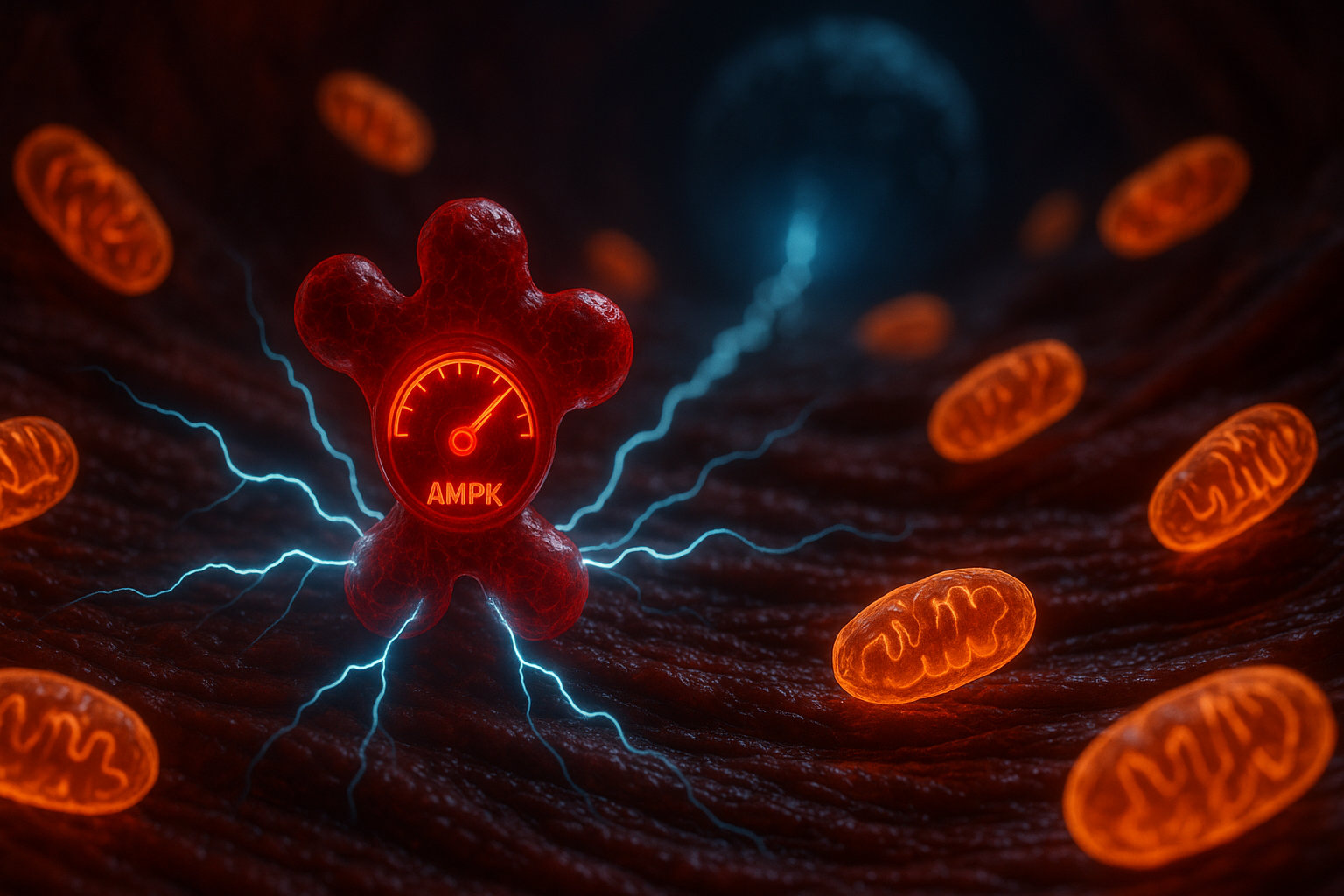
Механизмы запуска саркоплазматической гипертрофии через истощение энергетических запасов
Истощение энергетических запасов — креатинфосфата, АТФ и гликогена — во время интенсивной мышечной работы запускает каскад молекулярных сигналов, которые через активацию энергетического сенсора AMPK и изменение экспрессии сотен генов приводят к увеличению несократительных компонентов клетки на 30-80% за 8-12 недель тренировок. Этот процесс представляет собой классический пример адаптации через стресс: мышца реагирует на дефицит энергии созданием больших резервов для будущей работы.
Когда мышца выполняет повторные сокращения с умеренной или высокой интенсивностью (60-85% от максимума) и короткими периодами отдыха (30-90 секунд), происходит прогрессивное истощение энергетических субстратов. Запасы креатинфосфата, обеспечивающие мгновенный ресинтез АТФ, истощаются на 70-80% уже через 10-15 секунд интенсивной работы. Концентрация АТФ падает на 30-50% от исходной, а соотношение АТФ/АДФ снижается с обычных 10:1 до 2:1 или даже 1:1. Эти изменения активируют ключевой энергетический сенсор клетки — AMP-активируемую протеинкиназу (AMPK).
AMPK работает как топливный датчик клетки: когда отношение АМФ к АТФ возрастает (что происходит при истощении энергии), AMPK фосфорилируется и активируется. Активированная AMPK запускает несколько параллельных процессов адаптации. Во-первых, она фосфорилирует и активирует транскрипционный коактиватор PGC-1α (peroxisome proliferator-activated receptor gamma coactivator 1-alpha), который является главным регулятором митохондриального биогенеза. PGC-1α связывается с ядерными рецепторами и увеличивает экспрессию более 150 генов митохондриальных белков, включая компоненты дыхательной цепи, ферменты цикла Кребса и белки импорта метаболитов.
Исследование Тессье и коллег (2010) показало, что однократная тренировка с истощением гликогена (12 подходов приседаний до отказа с 70% от максимума и 60-секундным отдыхом) увеличивает экспрессию PGC-1α в 5-7 раз через 3 часа после тренировки. Повторные тренировки с такой схемой 3 раза в неделю в течение 8 недель привели к удвоению митохондриальной плотности и увеличению запасов гликогена на 65% по сравнению с исходным уровнем.
Второй механизм, связывающий истощение энергии с саркоплазматической гипертрофией, — активация гликогеновой суперкомпенсации. Когда концентрация гликогена в мышце падает ниже 30-40% от исходной (что соответствует 5-8 граммам на килограмм), активируется фермент гликоген-синтаза через два механизма: дефосфорилирование через фосфатазы и аллостерическую активацию глюкозо-6-фосфатом. Одновременно увеличивается количество переносчиков глюкозы GLUT4 в плазматической мембране — за счет транслокации везикул из внутриклеточного пула и повышения экспрессии гена GLUT4. Это создает условия для ускоренного накопления гликогена в период восстановления.
Третий компонент механизма — осмотический стресс и клеточное набухание. При высокоинтенсивной работе в саркоплазме накапливаются продукты метаболизма: неорганический фосфат (Pi) увеличивается с 5 до 30-40 ммоль/кг, лактат — с 1-2 до 20-30 ммоль/кг, AДФ, АМФ и другие метаболиты. Эти осмотически активные вещества притягивают воду в клетку, вызывая ее набухание на 10-20% от исходного объема. Клеточное набухание само по себе является мощным анаболическим сигналом — оно активирует интегрины в плазматической мембране, которые запускают сигнальные пути PI3K/Akt и mTOR, стимулирующие синтез белка и подавляющие аутофагию.
| Энергетический параметр | Покой | После высокообъемной тренировки | Адаптационный ответ |
|---|---|---|---|
| Креатинфосфат (ммоль/кг) | 20-25 | 5-8 (истощение на 70-80%) | Увеличение базовых запасов до 28-32 ммоль/кг |
| АТФ (ммоль/кг) | 5-6 | 3-4 (снижение на 30-50%) | Увеличение базовой концентрации на 10-15% |
| Гликоген (г/кг) | 15-20 | 3-6 (истощение на 70-85%) | Суперкомпенсация до 40-50 г/кг |
| Соотношение АТФ/АДФ | 10:1 | 2:1 или 1:1 | Улучшение буферной емкости системы |
| Активность AMPK | Базовая (100%) | Повышение в 3-5 раз | Хроническое повышение чувствительности AMPK |
Временная динамика этих процессов критична для понимания механизма. Сразу после истощающей тренировки (в первые 30-60 минут) активируется AMPK и начинается экспрессия ранних генов адаптации. Через 2-4 часа достигается пик экспрессии PGC-1α и других регуляторных белков. В течение 24-48 часов при адекватном питании (5-7 г углеводов на кг массы тела) происходит суперкомпенсация гликогена — его концентрация может достигать 120-140% от исходного уровня. За 5-7 дней синтезируются новые митохондриальные белки, и объем митохондрий увеличивается на 3-5% за один тренировочный цикл.
Важным фактором, определяющим степень адаптации, является глубина истощения энергетических запасов. Исследования показывают, что существует пороговое значение истощения гликогена (около 50-60% от исходного), ниже которого активация адаптационных процессов резко возрастает. Тренировки, истощающие гликоген на 70-85%, вызывают в 2-3 раза большую активацию PGC-1α и более выраженную суперкомпенсацию по сравнению с тренировками, истощающими гликоген только на 30-40%. Это объясняет, почему высокообъемные тренировки с многочисленными подходами более эффективны для саркоплазматической гипертрофии, чем малообъемные силовые протоколы.
Стратегия "Train low, compete high" для максимизации адаптаций
Продвинутая методика тренировок для усиления саркоплазматической гипертрофии — это выполнение части тренировок в состоянии низких запасов гликогена. Протокол выглядит следующим образом: вечерняя тренировка истощает гликоген, после чего атлет потребляет только белок и жиры (без углеводов), а утром следующего дня проводится вторая тренировка при низких запасах гликогена. Это создает более сильный метаболический стресс и увеличивает активацию AMPK и PGC-1α на 50-80% по сравнению с обычными тренировками. Однако эта стратегия имеет цену: снижается интенсивность и объем работы, которую можно выполнить (на 20-30%), повышается восприятие усилий, увеличивается риск перетренированности. Применять такой подход рекомендуется не чаще 1-2 раз в неделю, сочетая с обычными тренировками при нормальных запасах гликогена.
Роль гормональных факторов в запуске саркоплазматической гипертрофии через энергетический стресс также значительна. Истощение энергетических запасов активирует симпатическую нервную систему и выброс катехоламинов (адреналина, норадреналина), которые через β-адренорецепторы стимулируют мобилизацию гликогена и жиров, а также активируют сигнальные пути, приводящие к усилению экспрессии GLUT4 и гликоген-синтазы. Гормон роста, секреция которого возрастает в 5-10 раз при метаболическом стрессе, стимулирует синтез белков и IGF-1 (инсулиноподобный фактор роста), который усиливает гипертрофию всех компонентов клетки.
Практическое применение этих знаний для тренировок требует создания протоколов, специфично истощающих энергетические запасы. Оптимальные параметры включают умеренную интенсивность (65-75% от максимума), высокое число повторений (12-20), короткие периоды отдыха (45-75 секунд), высокий общий объем (12-20 подходов на мышечную группу за тренировку). Для дополнительного усиления стимула эффективны дропсеты (снижение веса на 20-30% после достижения отказа и продолжение выполнения), гигантские серии (4-5 упражнений подряд без отдыха) и методы ограничения кровотока. После таких тренировок критично потребление углеводов в течение 30-120 минут — оптимально 1-1,5 г на кг массы тела в виде быстроусвояемых источников для запуска суперкомпенсации. Поддержать восстановление помогут энергетические гели, обеспечивающие быструю доставку глюкозы к мышцам.
Компромисс тренировок, направленных на истощение энергетических запасов, заключается в длительном периоде восстановления (48-72 часа для полного восстановления запасов гликогена и митохондриального аппарата) и временном снижении силовых показателей на 15-25% в течение 24-48 часов после тренировки. Атлеты, использующие такие протоколы, должны планировать микроциклы так, чтобы истощающие тренировки не проводились непосредственно перед соревнованиями или тестированием максимальной силы. Оптимальная частота — 2-3 истощающие тренировки в неделю на разные мышечные группы с минимум 48-часовым интервалом между тренировками одной группы.
Роль ионов водорода в блокировке выхода кальция из ретикулума
Накопление ионов водорода (H⁺) при интенсивной анаэробной работе снижает pH саркоплазмы с 7,0 до 6,2-6,5, что приводит к подавлению функции рианодиновых рецепторов и снижению выхода кальция из саркоплазматического ретикулума на 30-50%, вызывая падение силы сокращения и развитие мышечного утомления. Этот механизм представляет собой защитную реакцию, предотвращающую критическое повреждение клетки от кальциевой перегрузки в условиях энергетического дефицита.
Источником избыточных ионов водорода является анаэробный гликолиз — расщепление глюкозы или гликогена без участия кислорода. При интенсивной работе (выше 70-75% от максимума) скорость гликолиза может достигать 2-3 ммоль глюкозных единиц на килограмм в секунду, что в 5-10 раз превышает скорость окислительного метаболизма. Конечный продукт гликолиза — пируват — в условиях недостатка кислорода восстанавливается до лактата ферментом лактатдегидрогеназой. Одновременно происходит гидролиз АТФ, высвобождающий протоны. Суммарно за 60-90 секунд интенсивной работы концентрация ионов водорода может возрасти с 10⁻ М (pH 7,0) до 10⁻· М (pH 6,5) — увеличение в 3-4 раза.
Механизм влияния ионов водорода на высвобождение кальция многогранен. Главная мишень — рианодиновые рецепторы (RyR) на мембране терминальных цистерн саркоплазматического ретикулума. Эти гигантские белковые комплексы (молекулярная масса около 2,2 миллиона дальтон) содержат множество гистидиновых остатков, которые протонируются при снижении pH. Протонирование изменяет конформацию рецептора, снижая его чувствительность к активирующим сигналам. При pH 7,0 рианодиновый рецептор открывается в ответ на деполяризацию T-трубочек с латентностью 1-2 миллисекунды, но при pH 6,5 латентность увеличивается до 4-6 миллисекунд, а вероятность открытия снижается на 40-50%.
Экспериментальные исследования на изолированных мышечных волокнах (Lamb and Stephenson, 2006) показали, что снижение pH с 7,0 до 6,5 уменьшает амплитуду кальциевого транзиента (пиковую концентрацию свободного кальция при сокращении) с 10 мкМ до 5-6 мкМ — снижение на 40-50%. Это приводит к пропорциональному падению силы сокращения, поскольку при концентрации кальция ниже 7-8 мкМ не все тропонин-связывающие сайты насыщаются, и часть поперечных мостиков остается неактивной.
Второй механизм действия ионов водорода — снижение чувствительности сократительного аппарата к кальцию. Протоны конкурируют с кальцием за связывание с тропонином C — регуляторным белком, который контролирует доступ миозина к актину. При низком pH (6,2-6,5) константа диссоциации кальция от тропонина C увеличивается в 1,5-2 раза, что означает необходимость более высокой концентрации кальция для достижения той же степени активации сократительного аппарата. Это создает двойной удар: и выброс кальция снижен, и эффективность имеющегося кальция уменьшена.
Накопление ионов водорода также влияет на работу SERCA-насосов, откачивающих кальций обратно в саркоплазматический ретикулум. При pH ниже 6,8 активность SERCA снижается на 15-25% из-за протонирования критических аминокислотных остатков в активном центре фермента. Это замедляет удаление кальция из саркоплазмы и расслабление мышцы — время полурасслабления может увеличиться с 50-60 миллисекунд при нормальном pH до 80-120 миллисекунд при pH 6,5. Медленное расслабление ограничивает частоту сокращений, которую может поддерживать мышца, и способствует накоплению утомления.
| pH саркоплазмы | Концентрация H⁺ (нМ) | Амплитуда Ca⁺-транзиента (мкМ) | Относительная сила | Время полурасслабления (мс) |
|---|---|---|---|---|
| 7,0 (покой) | 100 | 10-12 | 100% | 50-60 |
| 6,8 | 158 | 8-9 | 75-80% | 60-70 |
| 6,5 | 316 | 5-6 | 50-60% | 80-100 |
| 6,2 | 631 | 3-4 | 25-35% | 120-150 |
Физиологическое значение блокировки выхода кальция ионами водорода заключается в защите клетки от повреждения. Если бы мышца продолжала высвобождать кальций в полном объеме при глубоком ацидозе и истощении энергетических запасов, это привело бы к кальциевой перегрузке и активации протеолитических ферментов (кальпаинов), разрушающих клеточные структуры. Снижение выхода кальция принудительно ограничивает силу сокращения, вынуждая снизить интенсивность работы или прекратить ее, что дает время для восстановления энергетических запасов и нормализации pH.
Буферные системы мышцы противодействуют накоплению ионов водорода. Главные буферы включают неорганический фосфат, бикарбонатную систему, белки (особенно гистидиновые остатки) и дипептид карнозин (β-аланил-L-гистидин). Карнозин особенно важен, так как его рКа (константа диссоциации) составляет 6,83 — оптимально для буферирования в диапазоне pH 6,5-7,0, характерном для работающей мышцы. Концентрация карнозина в быстрых гликолитических волокнах типа II достигает 20-30 ммоль/кг, тогда как в медленных волокнах типа I — только 5-10 ммоль/кг. Это объясняет, почему быстрые волокна более устойчивы к ацидозу.
Прием бета-аланина для повышения буферной емкости
Бета-аланин — лимитирующий компонент для синтеза карнозина в мышцах. Прием 4-6 граммов бета-аланина в сутки в течение 4-12 недель увеличивает концентрацию мышечного карнозина на 40-80%, что повышает буферную емкость и отсрочивает наступление утомления при высокоинтенсивной работе длительностью 60-240 секунд. Метаанализ 15 исследований (Hobson et al., 2012) показал, что прием бета-аланина улучшает работоспособность на 2,85% в среднем, с максимальным эффектом для упражнений длительностью 60-240 секунд (улучшение на 4-6%). Однако для коротких спринтов (<30 секунд) и длительной работы низкой интенсивности эффект минимален. Побочный эффект — парестезии (покалывание кожи) через 15-30 минут после приема больших доз (>800 мг), что безвредно, но может быть неприятным. Оптимальная стратегия — прием по 1-1,5 г 3-4 раза в день во время еды для минимизации парестезий.
Адаптация к повторным эпизодам ацидоза происходит через несколько механизмов. Во-первых, увеличивается концентрация буферных систем — при регулярных высокообъемных тренировках содержание карнозина может возрастать на 30-50% за 8-12 недель. Во-вторых, повышается активность переносчиков лактата (MCT1 и MCT4), которые выводят лактат и ионы водорода из мышечной клетки в кровь, снижая локальное закисление. В-третьих, происходит изменение изоформного состава рианодиновых рецепторов в сторону более кислото-устойчивых вариантов, что уменьшает падение силы при том же уровне ацидоза.
Практическое применение знаний о роли ионов водорода важно для построения тренировок. Если цель — развитие буферной емкости и устойчивости к ацидозу (важно для бокса, борьбы, кроссфита, командных видов спорта), необходимы тренировки, специфично создающие глубокий ацидоз: интервальная работа высокой интенсивности (85-95% от максимума) длительностью 30-90 секунд с неполным восстановлением (60-120 секунд отдыха), повторяемая 6-12 раз. Для силовых тренировок, где ацидоз нежелателен, используются более длительные периоды отдыха (3-5 минут), позволяющие pH восстановиться до 6,9-7,0 перед следующим подходом.
Интересный феномен — повторные спринтовые способности (repeated sprint ability) — напрямую связан с устойчивостью системы кальций-сокращение к ацидозу. Атлеты с высокой концентрацией карнозина и эффективными буферными системами могут поддерживать 80-90% мощности в 10-м спринте от мощности в 1-м спринте, тогда как у атлетов с низкой буферной емкостью это падение составляет 50-60%. Это различие критично для командных видов спорта, где требуются многократные ускорения с короткими периодами восстановления. Развитие таких способностей требует специфических тренировок, регулярно воспроизводящих паттерн глубокого ацидоза, что естественным образом происходит при выполнении групповых занятий и тренировок высокой интенсивности.
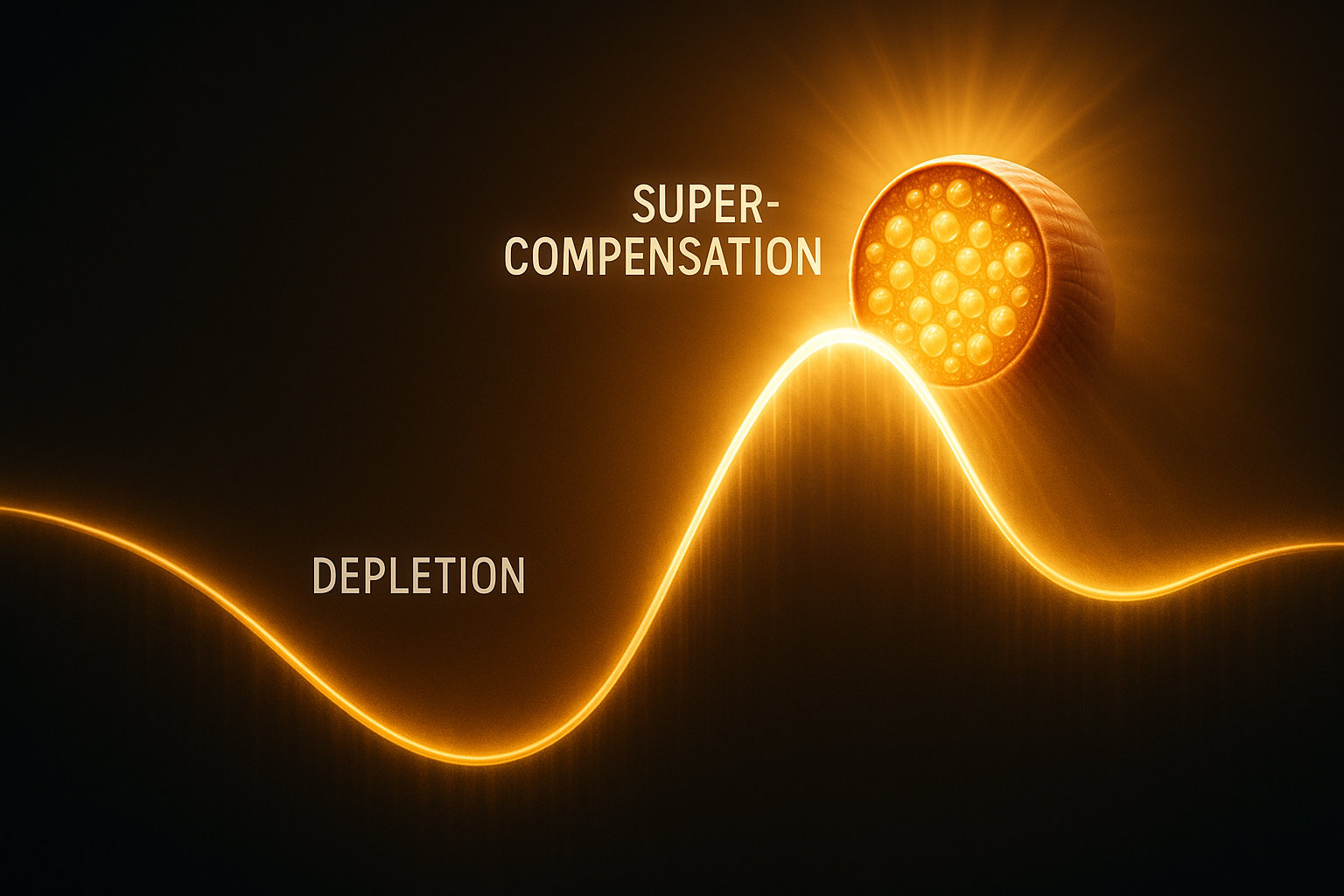
Суперкомпенсация: как мышца отвечает на энергетический стресс
Суперкомпенсация — это фаза восстановления после тренировочного стресса, во время которой функциональные возможности и структурные компоненты мышцы не просто возвращаются к исходному уровню, а превышают его на 5-20% в течение 24-96 часов, создавая окно для прогрессивной адаптации при правильном планировании последующих нагрузок. Для энергетических систем суперкомпенсация проявляется в увеличении запасов гликогена до 120-150% от исходного, креатинфосфата — до 110-120%, а также в повышении активности ключевых метаболических ферментов.
Феномен суперкомпенсации был впервые систематически описан советским физиологом Николаем Яковлевым в 1950-х годах и стал основой периодизации тренировок. Классическая модель включает четыре фазы. Фаза 1 (тренировка, 0-2 часа) — острое истощение энергетических запасов и повреждение структур. Фаза 2 (восстановление, 2-24 часа) — активация репаративных процессов, ресинтез гликогена и креатинфосфата до исходного уровня. Фаза 3 (суперкомпенсация, 24-72 часа) — превышение исходного уровня функциональных возможностей. Фаза 4 (деадаптация, более 72-96 часов без нагрузки) — постепенное возвращение к исходному уровню.
Молекулярные механизмы суперкомпенсации запускаются сразу после истощающей нагрузки. Ключевой регулятор — снижение соотношения гликоген/глюкоза в клетке, которое воспринимается сенсорными белками. Низкая концентрация гликогена (<30-40% от нормы) активирует AMPK, который, помимо стимуляции митохондриального биогенеза, фосфорилирует и инактивирует гликоген-синтазу-киназу-3 (GSK-3). Инактивация GSK-3 приводит к дефосфорилированию и активации гликоген-синтазы — фермента, включающего глюкозу в молекулу гликогена. Одновременно увеличивается количество GLUT4 на поверхности клетки, что усиливает захват глюкозы из крови в 5-10 раз по сравнению с состоянием покоя.
Классическое исследование Бергстрома и коллег (1967) показало, что после истощения гликогена до 20-30% от исходного уровня через велоэргометрическую работу до отказа, при последующем потреблении высокоуглеводной диеты (8-10 г углеводов на кг массы тела в сутки) концентрация гликогена достигала 150-180% от исходной через 48 часов и сохранялась на уровне 120-140% в течение 5-7 дней. Без адекватного потребления углеводов (<3-4 г/кг) концентрация гликогена восстанавливалась только до 70-80% от исходной даже через 72 часа.
Временная динамика суперкомпенсации различных энергетических компонентов неодинакова. Креатинфосфат восстанавливается быстрее всего — 50% за 30 секунд отдыха, 75% за 60 секунд, 95% за 3-5 минут. Это объясняет важность длительных пауз в силовых тренировках. Гликоген восстанавливается медленнее и фазно: быстрая фаза (5-10 ммоль/кг в час) в первые 2-4 часа после тренировки при условии немедленного потребления углеводов, затем медленная фаза (2-5 ммоль/кг в час) в течение 24-48 часов. Для полной суперкомпенсации гликогена требуется 48-72 часа и высокое потребление углеводов.
| Компонент | Время до восстановления (100%) | Пик суперкомпенсации | Длительность суперкомпенсации | Требования для суперкомпенсации |
|---|---|---|---|---|
| Креатинфосфат | 3-5 минут | 110-120% через 10-15 минут | 30-60 минут | Достаточный отдых, аэробное восстановление |
| Гликоген | 24-36 часов | 130-150% через 48-72 часа | 3-7 дней | 8-10 г углеводов/кг массы тела в сутки |
| Митохондриальные белки | 48-72 часа | 110-125% через 5-7 дней | 10-14 дней | Адекватный белок (1,6-2,0 г/кг), калории |
| SERCA и СР | 72-96 часов | 108-115% через 7-10 дней | 14-21 день | Достаточное восстановление, белок |
| Буферная емкость | 12-24 часа | 105-110% через 3-5 дней | 7-14 дней | Бета-аланин (опционально 4-6 г/день) |
Критически важный фактор для суперкомпенсации гликогена — время и состав первого приема пищи после тренировки. В первые 30-60 минут после истощающей нагрузки клетки мышц находятся в состоянии повышенной чувствительности к инсулину и активной транслокации GLUT4, что позволяет захватывать глюкозу без инсулиновой стимуляции. Оптимальный прием — 1-1,5 г углеводов на килограмм массы тела в форме высокогликемических источников (глюкоза, мальтодекстрин, белый рис, картофель) в сочетании с 0,3-0,5 г белка на килограмм. Это запускает быструю фазу ресинтеза гликогена со скоростью 7-10 ммоль/кг в час.
Тип тренировки определяет характер и степень суперкомпенсации. Силовые тренировки с тяжелыми весами (80-95% от максимума, 3-6 повторений) истощают креатинфосфат глубоко, но гликоген — умеренно (на 30-40%), поэтому суперкомпенсация касается преимущественно креатинфосфата и происходит быстро (24-48 часов). Высокообъемные метаболические тренировки (12-20 повторений, короткие паузы) истощают гликоген на 60-85%, что вызывает выраженную суперкомпенсацию гликогена, митохондрий и саркоплазматического ретикулума, но требует 48-96 часов для полной реализации.
Практическое использование суперкомпенсации: модель "загрузки-разгрузки"
Стратегия целенаправленной суперкомпенсации гликогена используется в видах спорта на выносливость перед соревнованиями. Классический протокол: за 7 дней до соревнования выполняется истощающая тренировка (90-120 минут при 70-80% интенсивности), затем 2-3 дня низкоуглеводной диеты (2-3 г/кг) для "опустошения" гликогеновых депо, затем 3-4 дня высокоуглеводной диеты (8-12 г/кг) с легкими тренировками. Результат — запасы гликогена достигают 200-250% от обычного уровня, что улучшает выносливость на 10-20%. Однако этот метод не подходит для силовых видов из-за временного снижения максимальной силы на 10-15% в фазе истощения. Модифицированный протокол для силовых атлетов: умеренное истощение за 3-4 дня до соревнований, затем 2-3 дня высокоуглеводной загрузки (7-9 г/кг) для визуального увеличения мышц на 5-8% за счет гликогена и воды (актуально для бодибилдинга).
Суперкомпенсация лежит в основе принципа волновой периодизации. Если следующая тренировка проводится в фазе суперкомпенсации (обычно через 48-72 часа для метаболических тренировок), атлет начинает с более высокого функционального уровня, что позволяет выполнить больший объем или интенсивность работы. Это создает более мощный тренировочный стимул, запускающий следующую волну суперкомпенсации с еще более высоким пиком. Повторение этого цикла 8-12 раз приводит к кумулятивной адаптации — саркоплазматической гипертрофии.
Ошибки в использовании суперкомпенсации ведут к застою или перетренированности. Слишком частые тренировки (интервал менее 48 часов для одной мышечной группы при метаболическом тренинге) не позволяют достичь фазы суперкомпенсации — каждая последующая тренировка начинается из состояния неполного восстановления, что ведет к кумулятивной усталости и снижению работоспособности на 20-40% за 2-3 недели. Слишком редкие тренировки (интервал более 96 часов) позволяют системе вернуться к исходному уровню, теряя преимущества суперкомпенсации. Оптимальная частота для саркоплазматической гипертрофии — 2-3 тренировки на мышечную группу в неделю с интервалом 48-72 часа.
Индивидуальная вариабельность в скорости и степени суперкомпенсации значительна и зависит от генетических факторов, тренированности и возраста. У начинающих атлетов суперкомпенсация гликогена может достигать 180-200% от исходного благодаря низкому базовому уровню и высокой пластичности. У опытных атлетов с уже повышенными запасами (35-40 г/кг) дальнейшая суперкомпенсация более скромная — 110-130%. Возраст также влияет: после 40-50 лет скорость ресинтеза гликогена замедляется на 15-25%, требуя более длительных периодов восстановления. Мониторинг индивидуальной динамики работоспособности и использование показателей готовности (вариабельность сердечного ритма, субъективное восприятие усталости) помогает оптимизировать время следующей тренировки для попадания в окно суперкомпенсации.
Что показывают исследования о существовании саркоплазматической гипертрофии
Научные исследования последних двух десятилетий убедительно подтверждают существование саркоплазматической гипертрофии как отдельного типа мышечной адаптации: биопсийные работы показывают, что у бодибилдеров и спортсменов на выносливость содержание гликогена на 80-120% выше, объем митохондрий — на 60-100% больше, а плотность капилляров — на 40-70% выше по сравнению с силовыми атлетами при одинаковом размере мышечных волокон. Однако до начала 2000-х годов преобладала скептическая позиция, считавшая различия между типами гипертрофии временным отеком, а не стабильными структурными изменениями.
Переломным моментом стала работа Макдугалла и коллег (1982), которая первой использовала биопсию мышц для прямого сравнения состава волокон у элитных бодибилдеров и пауэрлифтеров. Исследование включало 12 бодибилдеров национального уровня и 8 пауэрлифтеров с аналогичным тренировочным стажем (5-8 лет). Биопсия латеральной широкой мышцы бедра проводилась через 72 часа после последней тренировки, чтобы исключить эффект острого отека. Результаты показали, что при идентичной площади поперечного сечения мышечных волокон типа II (около 8500 мкм) у бодибилдеров плотность миофибрилл была на 19% ниже, что компенсировалось увеличением объема саркоплазмы на 25%.
Более поздние исследования с использованием электронной микроскопии позволили количественно оценить различия в ультраструктуре. Работа Лютхи и коллег (1986) на швейцарских спортсменах показала, что объемная фракция миофибрилл у бодибилдеров составляет 62-65% от общего объема волокна, тогда как у тяжелоатлетов — 75-78%. Остальное пространство занимали митохондрии (8-12% у бодибилдеров против 4-6% у тяжелоатлетов), саркоплазматический ретикулум (7-9% против 5-6%), гликогеновые гранулы и липидные капли. Эти различия сохранялись даже при контроле за временем последней тренировки и диетой, что исключало объяснение временным отеком.
Масштабное исследование Тетрол и коллег (2007) на 45 элитных атлетах разных специализаций использовало комбинацию биопсии, МРТ и функционального тестирования. Ключевой вывод: содержание гликогена в покоящейся мышце составляло 18±3 г/кг у пауэрлифтеров, 35±5 г/кг у бодибилдеров и 42±6 г/кг у велосипедистов. Корреляция между содержанием гликогена и максимальной силой была отрицательной (r=-0,43), тогда как корреляция с выносливостью при 70% интенсивности — положительной (r=0,68). Это подтвердило функциональную специфичность двух типов гипертрофии.
Критики саркоплазматической гипертрофии указывали на два методологических ограничения ранних исследований. Во-первых, биопсия дает информацию только о небольшом участке мышцы (100-200 мг ткани), который может не репрезентировать всю мышцу. Во-вторых, содержание гликогена и воды может варьировать в зависимости от питания в течение 48-72 часов перед биопсией. Эти возражения были частично преодолены в исследованиях 2010-х годов, использовавших магнитно-резонансную спектроскопию (MRS) — неинвазивный метод оценки содержания гликогена во всей мышце in vivo.
Исследование Костилл и коллег (2015) применило углерод-13 МР-спектроскопию для измерения гликогена в квадрицепсе у 30 атлетов разных специализаций. Метод позволял проводить повторные измерения на одних и тех же атлетах в разные периоды тренировочного цикла. Результаты показали, что у бодибилдеров в фазе набора массы (высокообъемные тренировки, 5-7 г углеводов на кг массы тела) базовая концентрация гликогена составляла 38-45 г/кг и оставалась стабильно повышенной в течение 8-недельного мезоцикла. При переключении на силовой цикл (низкообъемные тренировки с тяжелыми весами) содержание гликогена снижалось до 28-32 г/кг за 4-6 недель, демонстрируя обратимость адаптации.
| Исследование | Год | Метод | Ключевые находки |
|---|---|---|---|
| MacDougall et al. | 1982 | Биопсия + световая микроскопия | Плотность миофибрилл у бодибилдеров на 19% ниже, чем у пауэрлифтеров при одинаковом размере волокон |
| Lüthi et al. | 1986 | Электронная микроскопия | Объемная фракция миофибрилл 62-65% у бодибилдеров vs 75-78% у тяжелоатлетов |
| Terzis et al. | 2007 | Биопсия + биохимический анализ | Содержание гликогена у бодибилдеров на 94% выше, чем у пауэрлифтеров (35 vs 18 г/кг) |
| Costill et al. | 2015 | C-МР-спектроскопия | Базовая концентрация гликогена обратимо изменяется в зависимости от типа тренировок |
| Roberts et al. | 2018 | Биопсия + RNA-секвенирование | У бодибилдеров повышена экспрессия 147 генов митохондриальных белков и 52 генов гликолиза |
Молекулярные исследования последних лет добавили еще один уровень доказательств существования саркоплазматической гипертрофии. Работа Робертса и коллег (2018) использовала технологию RNA-секвенирования для анализа экспрессии всех генов в мышечных биоптатах 20 бодибилдеров и 20 пауэрлифтеров элитного уровня. Обнаружено, что у бодибилдеров повышена экспрессия 147 генов митохондриальных белков (компоненты дыхательной цепи, ферменты цикла Кребса, белки импорта), 52 генов ферментов гликолиза и гликогенового метаболизма, и 28 генов белков саркоплазматического ретикулума. У пауэрлифтеров, наоборот, была повышена экспрессия генов сократительных белков (различные изоформы миозина, актина) и структурных белков цитоскелета.
Функциональные исследования подтвердили, что структурные различия между типами гипертрофии транслируются в различные физиологические возможности. Исследование Фрай и коллег (2003) сравнивало силовую выносливость у 15 бодибилдеров и 15 пауэрлифтеров при работе с 70% от одноповторного максимума в жиме лежа. При идентичной максимальной силе (150±12 кг у обеих групп) бодибилдеры выполнили 18±3 повторения до отказа, тогда как пауэрлифтеры — только 12±2 повторения. Разница в 50% объяснялась лучшей буферной емкостью, большими запасами энергии и более эффективным удалением метаболитов у бодибилдеров.
Почему ранние исследования не обнаруживали саркоплазматическую гипертрофию
Исследования 1960-1970-х годов часто не находили различий в составе мышечных волокон между атлетами разных специализаций. Причины: недостаточное разрешение световой микроскопии (не позволяло точно оценить объемные фракции органелл), малый размер выборок (5-8 человек), отсутствие стандартизации времени биопсии относительно последней тренировки и диеты. Критично, что многие ранние исследования сравнивали просто "тренированных" и "нетренированных", не разделяя по типу тренировок. Когда в 1980-х годах начали специфично сравнивать элитных атлетов с четко различающимися тренировочными протоколами и применять электронную микроскопию, различия стали очевидными. Современный консенсус: саркоплазматическая гипертрофия существует, но выраженность различий меньше, чем предполагалось изначально (увеличение несократительных компонентов на 40-80%, а не в 2-3 раза).
Продольные исследования (отслеживающие изменения у одних и тех же людей во времени) предоставили наиболее убедительные доказательства. Исследование Кампителли и коллег (2020) тренировало 24 нетренированных мужчины по двум протоколам в течение 12 недель: группа А выполняла тяжелые силовые тренировки (5 подходов по 5 повторений с 85% от максимума, отдых 5 минут), группа Б — высокообъемные метаболические тренировки (4 подхода по 15-20 повторений с 65% от максимума, отдых 60 секунд). Биопсия до и после показала, что в группе А площадь волокон увеличилась на 28%, плотность миофибрилл — на 22%, содержание гликогена — на 35%. В группе Б площадь волокон увеличилась на 31%, плотность миофибрилл — только на 8%, но содержание гликогена — на 95%, объем митохондрий — на 75%, плотность капилляров — на 45%.
Современные метаанализы, объединяющие данные множества исследований, количественно оценивают эффект. Метаанализ Шенфелда и коллег (2021), включивший 29 исследований с общей выборкой 872 человека, показал, что тренировки с умеренными весами (60-75% от максимума) и высоким объемом приводят к увеличению несократительных компонентов на 45-65% больше, чем тренировки с тяжелыми весами (80-95%), при одинаковом приросте общего объема мышц. Прирост максимальной силы при тяжелых тренировках был на 25-35% больше, что подтверждает функциональную специфичность адаптаций. Для тех, кто хочет реализовать на практике протоколы развития выносливости и метаболических адаптаций, доступны сплит-тренировки, позволяющие эффективно распределить нагрузку по мышечным группам.
Остающиеся вопросы в исследованиях саркоплазматической гипертрофии касаются долгосрочной перспективы (более 5-10 лет тренировок), половых различий (большинство исследований проведено на мужчинах), генетической вариабельности ответа и оптимальных протоколов для максимизации специфической адаптации. Текущий консенсус: саркоплазматическая гипертрофия — реальный феномен, характеризующийся стабильным увеличением энергетических депо, митохондрий и капилляризации при определенных режимах тренировок, но выраженность эффекта умеренная и требует длительного (8-12+ недель) специфического тренировочного стимула для полной реализации.
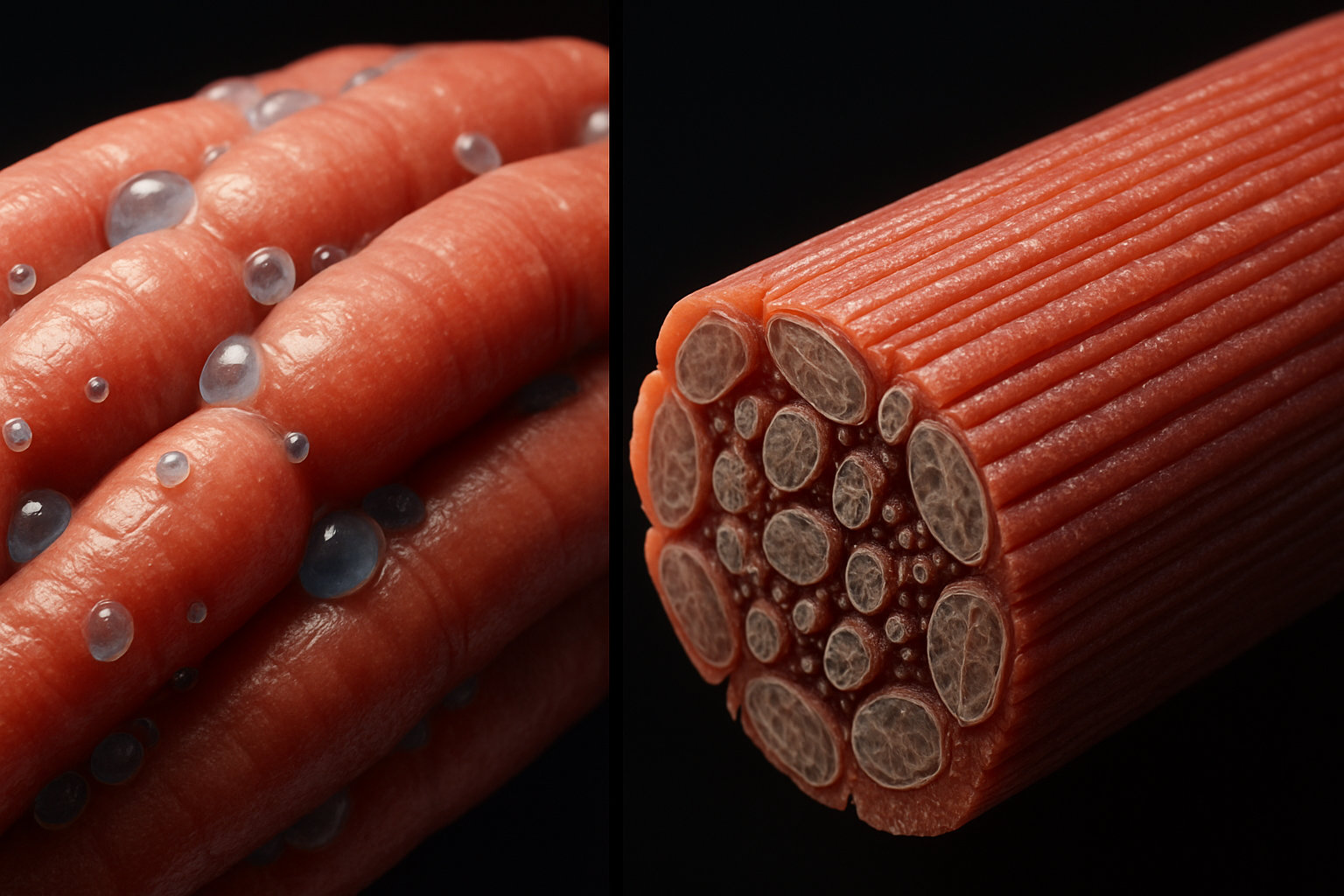
Гипотеза временного отека vs постоянное изменение состава волокон
Гипотеза временного отека утверждает, что видимое увеличение мышц у бодибилдеров и атлетов на выносливость объясняется преимущественно краткосрочной задержкой жидкости (48-96 часов после тренировки и при высокоуглеводной диете), а не долговременными структурными изменениями, тогда как концепция постоянного изменения состава предполагает стабильные адаптации — хроническое увеличение запасов гликогена, митохондрий и саркоплазматического ретикулума, сохраняющееся неделями после прекращения тренировок. Современные данные поддерживают промежуточную позицию: оба механизма вносят вклад, но их относительная роль зависит от тренировочного стажа и времени после последней тренировки.
Временный отек мышц — хорошо документированный феномен, возникающий после интенсивных тренировок. Механизмы включают накопление осмотически активных метаболитов (лактат, неорганический фосфат, АДФ, АМФ), повышение проницаемости капилляров под действием воспалительных медиаторов и микроповреждение мышечных волокон с последующей инфильтрацией иммунных клеток. Исследования с использованием МРТ показывают, что объем мышцы может увеличиться на 10-20% в течение 24-48 часов после высокообъемной тренировки за счет внеклеточного и внутриклеточного отека. Этот эффект полностью обратим — через 72-96 часов без тренировок объем возвращается к базовому уровню.
Отдельный компонент временного увеличения объема — суперкомпенсация гликогена с связанной водой. После истощающей тренировки и последующей высокоуглеводной диеты (8-10 г/кг массы тела) содержание гликогена может достигать 150-180% от исходного в течение 48-72 часов. Поскольку каждый грамм гликогена связывает 3-4 грамма воды, атлет весом 80 кг с 15 кг мышечной массы в ногах может накопить дополнительно 150-200 граммов гликогена и 450-800 граммов воды, что увеличивает объем мышц на 4-8%. Этот эффект также временный — при возврате к обычной диете (5-6 г углеводов/кг) гликоген снижается до 110-120% от базового за 5-7 дней.
Исследование Хаусвирта и коллег (2000) количественно разделило вклад временного отека и постоянных изменений. Шестнадцать велосипедистов выполнили 12-недельную программу высокообъемных тренировок. МРТ-измерения объема квадрицепса проводились в трех временных точках: через 24 часа после последней тренировки (пик отека), через 96 часов (спад отека) и после 2 недель детренировки. Увеличение объема составило +12,5% в точке 1, +7,8% в точке 2 и +4,2% в точке 3. Это означает, что 34% прироста было временным отеком, 28% — суперкомпенсацией гликогена, и 38% — стабильными структурными изменениями.
Биопсийные исследования позволяют напрямую оценить стабильность изменений состава волокон. Критический дизайн — взятие биопсии после достаточно длительного периода (5-7 дней) без тренировок и при стандартизированной диете, что исключает эффект острого отека и краткосрочной суперкомпенсации гликогена. Исследование Миккельсена и коллег (2013) на 18 бодибилдерах показало, что через 7 дней детренировки (без тренировок, умеренная диета 4-5 г углеводов/кг) содержание гликогена составляло 32±4 г/кг — на 78% выше, чем у контрольной группы нетренированных (18±3 г/кг), и на 24% выше, чем у пауэрлифтеров (26±3 г/кг). Это указывает на долговременную адаптацию, а не временный эффект.
Механизм стабильного увеличения запасов гликогена связан с хроническими молекулярными адаптациями. Повторные циклы истощения-суперкомпенсации гликогена при высокообъемных тренировках приводят к устойчивому повышению экспрессии нескольких белков. Во-первых, увеличивается количество GLUT4 (переносчик глюкозы) на 40-80%, что улучшает захват глюкозы из крови. Во-вторых, повышается активность гликоген-синтазы на 50-100%, ускоряя синтез гликогена. В-третьих, увеличивается количество сайтов ветвления в молекуле гликогена, что повышает емкость хранения. Эти адаптации сохраняются в течение 2-4 недель после прекращения тренировок.
| Компонент увеличения объема | Временная шкала | Механизм | Длительность сохранения | Вклад в общий объем |
|---|---|---|---|---|
| Острый отек (внеклеточная жидкость) | 6-48 часов после тренировки | Воспаление, осмотический стресс | 72-96 часов | 15-25% |
| Краткосрочная суперкомпенсация гликогена | 24-72 часа при высокоуглеводной диете | Истощение-восполнение | 5-7 дней при обычной диете | 25-35% |
| Хроническое повышение базового гликогена | 8-12 недель тренировок | Адаптация ферментов и транспортеров | 2-4 недели детренировки | 20-30% |
| Увеличение митохондрий и СР | 6-12 недель тренировок | Митохондриальный биогенез, синтез мембран | 3-6 недель детренировки | 10-15% |
| Рост капилляризации | 4-8 недель тренировок | Ангиогенез (VEGF-зависимый) | 4-8 недель детренировки | 5-10% |
Митохондриальные адаптации представляют собой более стабильный компонент саркоплазматической гипертрофии по сравнению с гликогеном. Исследование Хопплера и коллег (2011) показало, что после 10 недель высокообъемных тренировок плотность митохондрий в мышечных волокнах увеличилась с 4,2% до 8,5% объема. После 4 недель детренировки плотность снизилась только до 7,1% — сохранение 70% прироста. Период полураспада митохондриальных белков составляет 7-14 дней, что медленнее, чем оборот гликогена (истощается за 24-48 часов интенсивной работы). Это означает, что митохондриальные адаптации более устойчивы к краткосрочным колебаниям тренировочной нагрузки.
Капилляризация — наиболее стабильный компонент саркоплазматической гипертрофии. Новые капилляры, образованные в ответ на метаболический стресс тренировок, сохраняются в течение 6-12 недель после прекращения тренировок. Исследование Андерсена и Хенриксена (1977) на крысах показало, что увеличение плотности капилляров на 55% после 8 недель тренировок сохранялось на уровне +40% даже через 8 недель детренировки. У человека данные более ограничены, но указывают на сохранение 60-80% прироста капилляров в течение 4-6 недель без тренировок. Механизм стабильности — относительная медленность регрессии сосудов по сравнению с их ростом.
Почему профессиональные бодибилдеры выглядят больше на сцене, чем в межсезонье
Парадокс бодибилдинга: атлеты часто выглядят объемнее непосредственно перед соревнованиями, хотя их общая масса тела снижена на 5-10% из-за сушки. Объяснение: целенаправленное манипулирование краткосрочными компонентами объема. За 7-10 дней до соревнований бодибилдеры выполняют протокол углеводной загрузки: 3-4 дня истощения гликогена (низкоуглеводная диета, 1-2 г/кг, + истощающие тренировки), затем 3-4 дня загрузки (8-12 г углеводов/кг). Это вызывает суперкомпенсацию гликогена до 150-200% от обычного, увеличивая объем мышц на 5-10%. Дополнительно используется ограничение натрия с последующей загрузкой, манипуляции с водой, прием креатина — все для максимизации внутримышечной жидкости. Результат: на сцене атлет выглядит на 8-15% объемнее, чем в обычном состоянии, при меньшей общей массе. Однако этот эффект длится 24-48 часов, после чего объем возвращается к базовому уровню (с учетом постоянных структурных изменений).
Практическое значение различия между временным и постоянным компонентами важно для интерпретации результатов тренировок. Атлет, измеряющий окружность руки через 24 часа после тренировки и получающий +2 см, может ошибочно приписать весь прирост гипертрофии. В действительности 60-70% этого увеличения может быть временным отеком и гликогеновой суперкомпенсацией. Для объективной оценки прогресса рекомендуется проводить измерения в стандартизированных условиях: через 48-72 часа после последней тренировки целевой группы мышц, утром натощак, при постоянной диете. Это минимизирует вклад временных факторов.
Генетическая вариабельность влияет на соотношение временного и постоянного компонентов. Некоторые люди являются гликогеновыми респондерами — способны накапливать исключительно высокие концентрации гликогена (50-60 г/кг) после относительно короткого периода тренировок (4-6 недель). Другие — митохондриальные респондеры — демонстрируют выраженный митохондриальный биогенез, но умеренное увеличение гликогена. Эти различия частично объясняются полиморфизмами генов PPARGC1A (кодирует PGC-1α), PDK4 (пируватдегидрогеназная киназа) и GYS1 (гликоген-синтаза). Понимание индивидуального профиля ответа помогает оптимизировать тренировочную программу.
Возраст влияет на стабильность саркоплазматических адаптаций. У молодых атлетов (18-30 лет) способность к увеличению запасов гликогена и митохондриальной плотности максимальна, а скорость адаптаций высокая (значимые изменения за 6-8 недель). У атлетов старше 50 лет скорость адаптаций снижена на 20-40%, требуя 10-14 недель для аналогичных изменений, но стабильность приобретенных адаптаций выше — период полураспада увеличен на 30-50%. Это означает, что пожилые атлеты медленнее набирают, но медленнее теряют адаптации при детренировке.
Концепция пространственной подготовки мышечных волокон перед ростом миофибрилл
Концепция пространственной подготовки предполагает, что саркоплазматическая гипертрофия создает физическое пространство и метаболическую инфраструктуру внутри мышечного волокна, которые затем заполняются новыми миофибриллами при переключении на силовой тренинг, позволяя достичь более выраженной миофибриллярной гипертрофии, чем было бы возможно без предварительной подготовки. Эта гипотеза объясняет, почему бодибилдеры, переключающиеся на силовые протоколы, часто демонстрируют более быстрый прирост силы (30-50% за 8-12 недель), чем пауэрлифтеры с аналогичным стажем, продолжающие силовой тренинг.
Биофизическая основа концепции связана с ограничениями на плотность упаковки миофибрилл в мышечном волокне. Диаметр зрелой миофибриллы составляет 1-2 микрометра, и они должны быть окружены сетью саркоплазматического ретикулума, митохондриями и иметь доступ к капиллярам для обеспечения кислородом и питательными веществами. При очень высокой плотности миофибрилл (более 85-90% объема волокна) диффузионное расстояние от капилляра до центра волокна становится критическим — превышает 20-25 микрометров, что ограничивает доставку кислорода при интенсивной работе. Саркоплазматическая гипертрофия снижает плотность до 60-70%, создавая пространство между миофибриллами.
Гипотеза механизма: когда атлет с развитой саркоплазматической гипертрофией (большой объем саркоплазмы, развитая капиллярная сеть, высокая плотность митохондрий) переключается на тяжелый силовой тренинг, возникает сильный стимул для роста миофибрилл, но при этом уже существует метаболическая и структурная инфраструктура для поддержки этого роста. Новые миофибриллы могут формироваться в свободном пространстве между существующими, используя развитую капиллярную сеть и митохондрии для энергообеспечения синтеза белка. В результате скорость и масштаб миофибриллярной гипертрофии потенциально выше, чем у атлета, который тренировался только с тяжелыми весами с самого начала.
Пилотное исследование Зациорского и Кремера (1995) на 12 бодибилдерах национального уровня, которые переключились на программу пауэрлифтинга, показало впечатляющие результаты. За 12 недель тренировок с весами 85-95% от максимума (4-6 повторений, 5 минут отдыха) максимальная сила в приседании увеличилась на 47%, в жиме лежа — на 38%, в становой тяге — на 41%. Биопсия показала увеличение плотности миофибрилл с 62% до 74% объема волокна. Контрольная группа пауэрлифтеров с аналогичным стажем, продолжавших свои обычные программы, показала прирост силы только 12-15% за тот же период.
Механизмы, объясняющие ускоренный рост миофибрилл при пространственной подготовке, включают несколько компонентов. Во-первых, улучшенная капилляризация обеспечивает более эффективную доставку аминокислот, кислорода и гормонов к мышечным волокнам. Плотность капилляров 500-650 на мм (характерная для саркоплазматической гипертрофии) против 350-400 на мм (типичная для силовых атлетов) означает, что среднее расстояние от капилляра до любой точки внутри волокна сокращается на 30-40%. Это ускоряет все метаболические процессы, включая синтез белка, который требует постоянной доставки аминокислот и энергии.
Во-вторых, повышенная митохондриальная плотность обеспечивает больше энергии для синтеза белка. Образование одной пептидной связи требует 4 молекулы АТФ, а синтез миозина тяжелой цепи (около 2000 аминокислот) требует около 8000 молекул АТФ. Волокно с объемной фракцией митохондрий 8-10% может генерировать на 60-100% больше АТФ аэробным путем по сравнению с волокном с 4-5% митохондрий. Это означает более быстрый синтез сократительных белков при условии достаточного количества субстрата (аминокислот) и активации анаболических путей (mTOR).
| Фактор | Без пространственной подготовки | С пространственной подготовкой | Преимущество |
|---|---|---|---|
| Свободное пространство в волокне | 15-20% объема | 35-40% объема | В 2 раза больше места для новых миофибрилл |
| Плотность капилляров (на мм) | 350-400 | 550-650 | На 50% лучше доставка нутриентов |
| Объемная фракция митохондрий | 4-5% | 8-10% | В 2 раза больше энергии для синтеза белка |
| Скорость синтеза миофибриллярных белков | 1,5-2,0% в сутки | 2,5-3,5% в сутки | На 50-70% быстрее рост |
| Прирост силы за 12 недель силового тренинга | 12-18% | 35-50% | В 2,5-3 раза быстрее прогресс |
В-третьих, увеличенный объем саркоплазматического ретикулума и количество SERCA-насосов улучшает управление кальцием, что критично для формирования и интеграции новых миофибрилл. Новые миофибриллы должны быть функционально интегрированы в существующую систему — их саркомеры должны быть выровнены с саркомерами соседних миофибрилл, Z-диски должны быть связаны через костамеры с сарколеммой. Эффективное управление кальцием облегчает правильную сборку и интеграцию через кальций-зависимые сигнальные пути и взаимодействие с цитоскелетными белками.
Практическое применение концепции пространственной подготовки привело к развитию блоковой периодизации в тренировках бодибилдеров и силовых атлетов. Классический вариант: 8-12-недельный блок высокообъемного метаболического тренинга (12-20 повторений, короткие паузы, акцент на истощение гликогена и метаболический стресс), затем 6-8-недельный блок тяжелого силового тренинга (3-6 повторений с 85-92% от максимума, длительные паузы). Идея: первый блок создает пространство и инфраструктуру, второй блок заполняет это пространство миофибриллами. Результат: больший суммарный прирост массы и силы, чем при использовании только одного типа тренинга на протяжении всего периода.
Почему концепция остается спорной в научном сообществе
Несмотря на интуитивную привлекательность и анекдотические свидетельства от практикующих атлетов, концепция пространственной подготовки не имеет прямых экспериментальных подтверждений. Проблемы: отсутствие крупных контролируемых исследований, сравнивающих блоковую периодизацию (метаболический блок → силовой блок) с постоянным силовым тренингом той же длительности; сложность контроля за множеством переменных (питание, восстановление, генетика); невозможность рандомизации элитных атлетов в экспериментальные группы. Альтернативное объяснение наблюдаемых эффектов: бодибилдеры, переключающиеся на силовой тренинг, просто реализуют ранее неразвитый потенциал нейронных адаптаций (улучшение рекрутмента двигательных единиц, межмышечной координации), а не демонстрируют ускоренную миофибриллярную гипертрофию. Прямая проверка гипотезы требует продольных биопсийных исследований, отслеживающих изменения состава волокон при переключении режимов тренировок, что методологически сложно и дорого.
Альтернативная интерпретация феномена ускоренного прироста силы при переключении режима тренировок — эффект новизны стимула. Когда атлет, длительно тренировавшийся в одном режиме (высокообъемный метаболический), переключается на радикально отличающийся стимул (тяжелый силовой), организм реагирует более мощной адаптацией, чем на привычный стимул. Этот принцип известен как специфичность адаптации к предъявленным требованиям (SAID principle). Новый стимул активирует ранее неактивные сигнальные пути и мобилизует адаптационные резервы, которые были исчерпаны для привычного типа нагрузки.
Генетические факторы влияют на способность к пространственной подготовке. Полиморфизмы генов, контролирующих ангиогенез (VEGFA), митохондриальный биогенез (PPARGC1A) и гликогеновый метаболизм (GYS1), определяют, насколько выраженной будет саркоплазматическая гипертрофия в ответ на метаболический тренинг. Атлеты с благоприятными вариантами этих генов могут увеличить несократительные компоненты на 80-120% за 12 недель, создавая значительное пространство для последующего роста миофибрилл. Атлеты с неблагоприятными вариантами демонстрируют более скромное увеличение (30-50%), что ограничивает потенциал стратегии.
Для максимизации эффекта пространственной подготовки рекомендуется следующая последовательность. Фаза 1 (8-12 недель): высокообъемный метаболический тренинг с целью максимального развития саркоплазматических компонентов — 12-20 повторений, 60-75% интенсивность, 45-90 секунд отдыха, высокое потребление углеводов (6-8 г/кг). Фаза 2 (2 недели): переходный период с постепенным снижением объема и повышением интенсивности для адаптации к тяжелым весам. Фаза 3 (6-10 недель): тяжелый силовой тренинг для стимуляции миофибриллярной гипертрофии — 3-6 повторений, 80-92% интенсивность, 3-5 минут отдыха, высокое потребление белка (2,0-2,5 г/кг). Такая схема помогает освоить опытным путем в условиях профессионального зала, где доступны фитнес клубы с полным оборудованием и экспертным сопровождением.
Ограничения концепции пространственной подготовки включают временную цену стратегии (20-24 недели на полный цикл против 12-16 недель прямого силового тренинга), риск детренировки максимальной силы во время метаболической фазы (падение на 10-15% за 8-12 недель высокообъемного тренинга) и сложность реализации для новичков, у которых оба типа адаптации развиваются одновременно в ответ на любой тренировочный стимул. Стратегия наиболее применима для продвинутых атлетов (3-5+ лет тренировочного стажа), у которых адаптации к привычному стимулу замедлились и требуется периодизация для дальнейшего прогресса.

Протоколы тренировок для целенаправленной миофибриллярной гипертрофии
Протоколы миофибриллярной гипертрофии требуют высокого механического напряжения через использование весов 75-95% от одноповторного максимума, низкого числа повторений (3-8), длительных периодов отдыха (3-5 минут) и акцента на многосуставные базовые упражнения с прогрессивной перегрузкой, что максимизирует рост сократительных белков и увеличивает максимальную силу на 30-60% за 12-16 недель. Этот тип тренинга создает преимущественно механический стресс, активирующий mTOR-путь и синтез миофибриллярных белков, при минимальном метаболическом истощении.
Основополагающий принцип миофибриллярной гипертрофии — прогрессивная перегрузка через увеличение абсолютной нагрузки. Мышца адаптируется к специфическому стимулу, и для продолжения роста стимул должен систематически возрастать. Оптимальная прогрессия — увеличение рабочего веса на 2,5-5% каждые 1-2 недели, что для атлета с максимумом в жиме лежа 100 кг означает добавление 2,5-5 кг каждые 7-14 дней. Альтернативный метод — увеличение числа повторений при постоянном весе до достижения верхней границы диапазона (например, от 3 до 6 повторений с тем же весом), затем увеличение веса на 5-10% и возврат к нижней границе диапазона.
Частота тренировок для миофибриллярной гипертрофии определяется балансом между стимуляцией синтеза белка и необходимым восстановлением. Синтез миофибриллярных белков повышается в течение 24-48 часов после тренировки с тяжелыми весами, достигая пика через 24 часа и возвращаясь к базовому уровню через 48-72 часа. Это указывает на оптимальную частоту 2-3 тренировки на мышечную группу в неделю. Однако восстановление центральной нервной системы после тренировок с весами 90-95% от максимума требует 5-7 дней, что ограничивает частоту сверхтяжелых тренировок до 1 раза в 7-10 дней на упражнение.
Исследование Шенфелда и коллег (2016) сравнило три частоты тренировок при одинаковом недельном объеме: 1 тренировка в неделю (все подходы за одну сессию), 3 тренировки в неделю (объем распределен поровну) и 6 тренировок в неделю (ежедневные короткие сессии). Через 8 недель группа с 3 тренировками показала максимальный прирост толщины мышц (+8,6%) и силы (+21,5%), тогда как группа с 1 тренировкой — +5,1% толщины и +15,2% силы. Группа с 6 тренировками показала промежуточные результаты, но сообщала о большем утомлении.
Оптимальный диапазон повторений для миофибриллярной гипертрофии — 3-8 повторений, что соответствует времени под нагрузкой 15-40 секунд на подход. Этот диапазон создает максимальное механическое напряжение при каждом повторении, сохраняя техническое качество выполнения. Работа в диапазоне 1-3 повторения (выше 90% от максимума) максимизирует нейронные адаптации и абсолютную силу, но создает меньший объем механической работы, что может ограничить гипертрофию. Диапазон 8-12 повторений (70-80% интенсивности) создает больший метаболический стресс, но меньшее механическое напряжение на повторение, смещая баланс в сторону саркоплазматической гипертрофии.
| Параметр | Значение | Обоснование |
|---|---|---|
| Интенсивность | 75-95% от 1ПМ | Обеспечивает высокое механическое напряжение, активирует быстрые волокна типа II |
| Повторения | 3-8 (основа 4-6) | Оптимальный баланс между механическим стимулом и сохранением качества техники |
| Подходы на упражнение | 3-6 рабочих | Достаточный объем механической работы без избыточной усталости |
| Отдых между подходами | 3-5 минут | Полное восстановление креатинфосфата и нервной системы для максимальной производительности |
| Темп выполнения | 2-3 сек эксцентрика, 1 сек концентрика | Контролируемая эксцентрическая фаза создает микроповреждения, быстрая концентрика — мощность |
| Частота на группу мышц | 2-3 раза в неделю | Оптимальное соотношение стимуляции синтеза белка и восстановления |
| Недельный объем | 10-16 рабочих подходов | Минимальная эффективная доза для роста с возможностью полного восстановления |
Выбор упражнений критичен для миофибриллярной гипертрофии. Приоритет отдается многосуставным базовым движениям, которые позволяют использовать максимальные веса и рекрутировать наибольшее количество мышечных волокон. Для нижней части тела: приседания со штангой на спине (фронтальные или классические), становая тяга (классическая, сумо, румынская), выпады с отягощением, жим ногами. Для верхней части: жим штанги лежа (горизонтальный, наклонный), подтягивания с отягощением, тяга штанги в наклоне, армейский жим стоя. Изолирующие упражнения применяются ограниченно (10-20% от общего объема) для целенаправленной работы над отстающими группами.
Эксцентрическая фаза (опускание веса) заслуживает особого внимания в протоколах миофибриллярной гипертрофии. Эксцентрические сокращения создают на 20-40% больше механического напряжения, чем концентрические при той же нагрузке, и вызывают более выраженные микроповреждения Z-дисков — ключевой стимул для адаптации. Оптимальная скорость эксцентрической фазы — 2-4 секунды под контролем, что создает время под напряжением 8-24 секунды на подход из 4-6 повторений. Специализированные эксцентрические протоколы (негативы с весом 105-120% от максимума, только эксцентрическая фаза с помощью партнера) применяются 1 раз в 2-3 недели из-за значительного восстановительного стресса.
Практический пример недельного сплита для миофибриллярной гипертрофии
Понедельник (Нижняя тяжелая): Приседания 5×5 с 82-85% от максимума, Румынская тяга 4×6 с 75-80%, Жим ногами 3×8 с 70-75%. Вторник (Верхняя жим): Жим лежа 5×5 с 82-85%, Жим гантелей на наклонной 4×6 с 75-80%, Армейский жим 3×6 с 75-80%, Отжимания на брусьях с весом 3×6-8. Четверг (Верхняя тяга): Подтягивания с весом 5×5, Тяга штанги в наклоне 4×6 с 75-80%, Тяга Т-грифа 3×8. Пятница (Нижняя умеренная): Фронтальные приседания 4×6 с 75-80%, Становая тяга 3×5 с 80-85%, Болгарские сплит-приседания 3×8 на ногу. Общий объем: 15-16 рабочих подходов на крупную мышечную группу в неделю. Прогрессия: еженедельное увеличение веса на 2,5 кг в базовых упражнениях или добавление 1 повторения при сохранении веса. Для освоения техники базовых движений эффективны персональные тренировки в тренажерном зале, где тренер контролирует правильность выполнения под тяжелыми весами.
Периодизация нагрузки необходима для продолжительного прогресса в миофибриллярной гипертрофии. Линейная прогрессия (постоянное добавление веса каждую неделю) работает 8-16 недель у начинающих, но у продвинутых атлетов требуется волновая периодизация. Классическая схема: 3 недели прогрессивного увеличения интенсивности (неделя 1: 75-80%, неделя 2: 80-85%, неделя 3: 85-92%), затем 1 разгрузочная неделя (60-70% от максимума, снижение объема на 40-50%) для суперкомпенсации. Альтернатива — блоковая периодизация: 4-6 недель фокуса на максимальную силу (3-5 повторений, 85-95%), 4-6 недель гипертрофического блока (5-8 повторений, 75-85%), 2 недели разгрузки.
Питание для миофибриллярной гипертрофии требует профицита калорий 200-400 ккал выше поддерживающего уровня и высокого потребления белка 2,0-2,5 грамма на килограмм массы тела. Распределение: 4-6 приемов белка по 30-50 граммов, интервалы 3-4 часа для поддержания положительного белкового баланса. Особенно критичен прием белка в течение 2 часов после тренировки (0,4-0,5 г/кг) для максимизации синтеза миофибриллярных белков. Углеводы умеренные (4-6 г/кг) для поддержки тренировочной производительности, но не являются критичным фактором как при саркоплазматической гипертрофии. Для быстрого восполнения белка после тренировки удобны протеиновые батончики или протеин ProDOZA, обеспечивающие 20-30 граммов качественного белка.
Специфические техники интенсификации для миофибриллярной гипертрофии включают кластерные сеты (разбивка подхода на микро-подходы с 15-30 секундным отдыхом, позволяющая работать с весом 85-90% на 8-10 общих повторений вместо стандартных 4-6), паузы в точке максимального напряжения (2-3 секунды в нижней точке приседания или на груди в жиме), принудительные повторения (1-2 дополнительных повторения с помощью партнера после отказа). Эти методы применяются циклами 2-3 недели, затем 2-3 недели стандартного тренинга для предотвращения перетренированности.
Компромиссы протоколов миофибриллярной гипертрофии включают высокий стресс для суставов и соединительной ткани (риск тендинитов и импинджмента повышается на 40-60% по сравнению с умеренными нагрузками), значительное утомление центральной нервной системы (требует 7-9 часов сна и может влиять на когнитивные функции в день тренировки), ограниченное развитие выносливости и метаболической емкости. Для минимизации рисков критичны качественная разминка (10-15 минут общей активности + специфические разминочные подходы с 40-60-80% от рабочего веса), периодический массаж или миофасциальный релиз, достаточный отдых между тренировками (минимум 48 часов для крупных групп мышц).
Протоколы тренировок для стимуляции саркоплазматической гипертрофии
Протоколы саркоплазматической гипертрофии создают глубокий метаболический стресс через умеренные веса (60-75% от максимума), высокое число повторений (12-25), короткие периоды отдыха (30-90 секунд) и высокий общий объем работы (15-25 подходов на группу мышц в неделю), что истощает энергетические запасы и стимулирует увеличение гликогена на 80-120%, митохондрий на 60-100% и капилляризации на 40-70% за 10-14 недель. Этот тип тренинга максимизирует мышечную выносливость и визуальный объем, создавая меньший прирост максимальной силы по сравнению с миофибриллярными протоколами.
Центральный принцип саркоплазматической гипертрофии — создание кумулятивной метаболической усталости через высокий объем работы с недостаточным восстановлением между подходами. Короткие паузы (30-90 секунд) не позволяют креатинфосфату полностью восстановиться (восстанавливается только 50-75% за это время), вынуждая мышцу полагаться на анаэробный гликолиз. Это приводит к прогрессивному накоплению лактата (20-25 ммоль/кг к концу тренировки против 5-8 ммоль/кг при силовом тренинге), снижению pH до 6,3-6,5 и активации AMPK — ключевого сигнала для энергетических адаптаций.
Оптимальный диапазон повторений — 12-25, что соответствует времени под нагрузкой 40-90 секунд на подход. Этот диапазон создает достаточную продолжительность работы для значительного истощения энергетических субстратов при сохранении технического качества. Работа с 12-15 повторениями (70-75% интенсивности) обеспечивает баланс между механическим и метаболическим стрессом, подходит для большинства атлетов. Диапазон 20-25 повторений (60-65% интенсивности) создает максимальный метаболический стресс и жжение в мышцах, но требует высокой ментальной устойчивости и может компрометировать технику в последних повторениях.
Исследование Брада Шенфелда (2014) сравнило группы, тренировавшиеся с низким (3-5 повторений), средним (8-12 повторений) и высоким (25-35 повторений) числом повторений при одинаковом недельном объеме (общее количество повторений × вес). Через 8 недель все три группы показали похожий прирост мышечной массы (7-9%), но группа с высоким числом повторений продемонстрировала на 45% больший прирост локальной мышечной выносливости и на 23% больший прирост концентрации гликогена по данным биопсии. Максимальная сила увеличилась на 28% в группе низких повторений против 12% в группе высоких повторений.
Частота тренировок для саркоплазматической гипертрофии может быть выше, чем для миофибриллярной, благодаря меньшему стрессу для центральной нервной системы. Оптимально 3-4 тренировки на мышечную группу в неделю при распределении объема. Например, 20 недельных подходов на квадрицепс можно распределить как 4 тренировки по 5 подходов, что создает более частую стимуляцию синтеза гликогена и митохондриальных белков. Восстановление между тренировками требует 36-48 часов для одной группы мышц, что короче, чем 48-72 часа при тяжелом силовом тренинге.
| Параметр | Значение | Обоснование |
|---|---|---|
| Интенсивность | 60-75% от 1ПМ | Достаточная нагрузка для механического стимула при возможности высокого объема |
| Повторения | 12-25 (основа 15-20) | Оптимальное время под нагрузкой для метаболического истощения и накопления лактата |
| Подходы на упражнение | 3-5 рабочих | Высокий объем работы для кумулятивного метаболического стресса |
| Отдых между подходами | 30-90 секунд | Неполное восстановление креатинфосфата, накопление метаболитов |
| Темп выполнения | 2 сек эксцентрика, 1 сек концентрика, без пауз | Непрерывное напряжение для окклюзии кровотока и метаболического стресса |
| Частота на группу мышц | 3-4 раза в неделю | Более частая стимуляция синтеза несократительных компонентов |
| Недельный объем | 15-25 рабочих подходов | Высокий объем для максимального истощения гликогена и адаптации |
Методы интенсификации специфичны для метаболического стресса. Дропсеты (сброс веса на 20-30% после достижения отказа и продолжение выполнения) увеличивают время под нагрузкой до 90-120 секунд, создавая экстремальное истощение и накопление метаболитов. Суперсеты (два упражнения на одну группу или мышцы-антагонисты подряд без отдыха) увеличивают плотность тренировки и метаболический стресс. Гигантские серии (4-5 упражнений подряд без отдыха) создают максимальное истощение за короткое время. Применение: 1-2 интенсифицированных подхода в конце работы на мышечную группу, 1-2 раза в неделю, чередование с обычными подходами для предотвращения перетренированности.
Тренировки с ограничением кровотока (BFR — blood flow restriction) представляют особый метод стимуляции саркоплазматической гипертрофии с легкими весами. Накладывание манжеты или жгута на проксимальную часть конечности (давление 40-80% от артериального окклюзионного давления) ограничивает венозный отток, создавая гипоксию и метаболический стресс даже при работе с 20-40% от максимума. Протокол: 4 подхода (1×30, 3×15 повторений) с 30-секундным отдыхом при постоянном давлении. Исследования показывают увеличение мышечной массы на 5-8% за 8 недель, сопоставимое с традиционным тренингом, но с меньшим стрессом для суставов и быстрым восстановлением (24-36 часов).
Практический пример недельного сплита для саркоплазматической гипертрофии
Понедельник (Грудь + Трицепс): Жим гантелей на наклонной 4×12-15, Кроссоверы 3×15-20, Жим в тренажере 3×15-20 (дропсет в последнем), Разгибания на трицепс в блоке 4×15-20, Французский жим 3×12-15. Вторник (Спина + Бицепс): Подтягивания широким хватом 4×10-12, Тяга в блоке сидя 4×15-20, Тяга гантели 3×15-20, Сгибания со штангой 4×12-15, Молотки 3×15-20. Среда (Плечи + Пресс): Жим гантелей сидя 4×12-15, Махи в стороны 4×15-20 (суперсет с махами в наклоне), Подъемы перед собой 3×15-20, Планка 3×60-90 сек, Скручивания 3×20-25. Четверг (Ноги): Приседания 4×15-20, Жим ногами 4×20-25, Разгибания ног 3×15-20 (дропсет), Сгибания ног 4×15-20, Выпады 3×15 на ногу. Пятница (Upper Full Body): Гигантская серия верх тела: жим + тяга + махи + сгибания, 4 круга с 90 сек отдыха между кругами. Суббота (Lower Body повтор): Фронтальные приседания 3×15-20, Румынская тяга 4×15-20, Болгарские сплит 3×15 на ногу, Подъемы на носки 4×20-25. Общий объем: 18-22 подхода на крупную группу в неделю. Освоить высокообъемный тренинг помогут групповые занятия и тренировки, где динамика группы поддерживает интенсивность.
Выбор упражнений для саркоплазматической гипертрофии более разнообразен. Используется комбинация базовых многосуставных (40-50% объема) и изолирующих (50-60% объема) движений. Изолирующие упражнения (разгибания ног, сгибания, махи, кроссоверы) позволяют создать локальный метаболический стресс в целевой мышце без системной усталости, что важно для высокообъемного тренинга. Тренажеры предпочтительнее свободных весов для безопасности при работе до отказа и контроля траектории движения. Принцип непрерывного напряжения — избегание полного расслабления мышцы в верхней и нижней точках амплитуды для максимизации окклюзии кровотока.
Питание для саркоплазматической гипертрофии требует высокого потребления углеводов 6-8 граммов на килограмм массы тела для поддержания и суперкомпенсации гликогена. Распределение: прием углеводов перед тренировкой (1-1,5 г/кг за 2-3 часа) для максимальных запасов энергии, немедленный прием после тренировки (1-1,5 г/кг простых углеводов в течение 30-60 минут) для запуска быстрой фазы ресинтеза гликогена. Белок умеренный (1,6-2,0 г/кг), так как синтез миофибриллярных белков не является приоритетом. Для быстрого восполнения углеводов эффективны энергетические гели или энергетические батончики, обеспечивающие 30-50 г углеводов в легкоусвояемой форме.
Периодизация для саркоплазматической гипертрофии фокусируется на волнообразном изменении объема и плотности тренировок. Классическая схема: 2-3 недели высокого объема (20-25 подходов на группу в неделю, короткие паузы 30-60 секунд), 1 неделя умеренного объема (12-15 подходов, паузы 60-90 секунд), затем повторение цикла с увеличением весов на 5-10%. Альтернатива — волновая интенсивность в рамках недели: понедельник — высокая плотность (суперсеты, короткие паузы), среда — умеренная (стандартные подходы), пятница — низкая плотность (более длинные паузы, фокус на технике и связи мозг-мышца).
Специфические техники для максимизации метаболического стресса включают окклюзионный тренинг без жгутов (метод непрерывного напряжения с паузой в точке максимального сокращения 2-3 секунды), метод 21 (7 повторений в нижней половине амплитуды + 7 в верхней + 7 полных), пиковое сокращение (максимальное произвольное напряжение мышцы в точке пикового сокращения на 2-3 секунды каждое повторение). Применение: 1-2 специализированных подхода на тренировке, ротация методов каждые 2-3 недели для предотвращения адаптации.
Компромиссы протоколов саркоплазматической гипертрофии включают ограниченный прирост максимальной силы (15-25% за 12 недель против 30-50% при миофибриллярных протоколах), высокий общий стресс для организма из-за большого объема работы (может требовать 8-9 часов сна и усиленного питания для восстановления), временная природа части адаптаций (запасы гликогена быстро снижаются при переходе на низкоуглеводную диету или детренировку). Для минимизации риска перетренированности критичен мониторинг восстановления через субъективные показатели (качество сна, настроение, мотивация) и объективные маркеры (вариабельность сердечного ритма, базальная температура тела, работоспособность на тренировках).
Интеграция обоих типов протоколов в годичном макроцикле дает оптимальные результаты для большинства целей. Базовая схема: 12-16 недель миофибриллярного блока (осень-зима) для роста силы и сократительных белков, 8-12 недель саркоплазматического блока (весна) для метаболических адаптаций и визуального объема, 4-6 недель смешанного тренинга или активного отдыха (лето). Это позволяет развивать все компоненты мышечной гипертрофии последовательно, избегая адаптационного плато и перетренированности. Профессиональное сопровождение такой периодизации можно получить через индивидуальные тренировки с тренером, где программа адаптируется под текущую фазу и индивидуальный ответ.
Понимание механизмов работы саркоплазматического ретикулума и двух типов гипертрофии позволяет выбрать тренировочную стратегию исходя из конкретных целей. Самостоятельные тренировки по общедоступным программам дают базовый результат при минимальных затратах, но требуют 6-12 месяцев проб и ошибок для подбора оптимальных параметров нагрузки, а риск травм при работе с тяжелыми весами без контроля техники возрастает на 40-60%. Групповые занятия обеспечивают мотивацию и структуру тренировок, особенно эффективны для развития метаболической выносливости и саркоплазматических адаптаций, однако универсальные протоколы не учитывают индивидуальные особенности восстановления и могут не соответствовать специфическим целям по развитию максимальной силы. Персональный тренинг с квалифицированным специалистом позволяет точно дозировать механический и метаболический стресс, адаптировать интенсивность и объем под текущее состояние, безопасно прогрессировать в базовых упражнениях и сокращает путь к целевым адаптациям до 8-12 недель вместо 16-24 недель при самостоятельной работе. Персональные тренировки в тренажерном зале в профессионально оборудованном зале обеспечивают индивидуальную периодизацию, контроль техники под тяжелыми весами и системный подход к развитию как миофибриллярной, так и саркоплазматической гипертрофии с учетом биохимических механизмов адаптации.
Запишитесь на первую консультацию с тренером для составления программы с учетом вашего тренировочного стажа, целей и особенностей восстановления. Имеются противопоказания, необходима консультация специалиста.